УДК 575.174
Калмыки и генетическое наследие экспансии тюркоязычных кочевников в западной Евразии
А.М. Тюрин
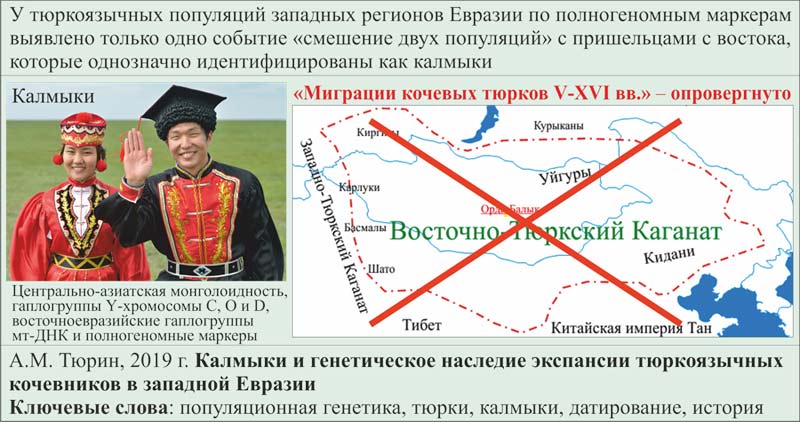
Ключевые слова: популяционная генетика, тюрки, калмыки, датирование, история.
Введение
С точки зрения популяционной генетики, тюрки – это разные популяции, говорящие на диалектах тюркского языка. Авторы публикации [Yunusbayev et al., 2015] поставили вполне определенные исследовательские задачи, сформулированные в виде вопросов:
1. Имеется ли в генофонде современных тюрок общий сигнал (определенная совокупность генетических маркеров)?
2. Связан ли он с доисторическими кочевыми («prehistoric nomadic») тюрками?
3. Есть ли общее для всех тюрков наследие, которое можно проследить до их гипотетической внутриазитской родины («Inner Asian Homeland», территории Монголии, Южной Сибири и Северного Китая)?
4. Можно ли подтвердить по данным популяционной генетики на основе применения соответствующего инструментария общее происхождение тюркоязычных народов, локализованных сегодня в Западной Евразии и во внутриазитской родине?
В качестве рабочей гипотезы приято следующее. «The earliest Turkic ruled polities (between the 6th and 9th centuries) were centered in what is now Mongolia, northern China, and southern Siberia. Accordingly, this region has been put forward as the point of origin for the dispersal of Turkic-speaking pastoral nomads. We designate it here as an «Inner Asian Homeland»».
Для поиска ответов на поставленные вопросы выполнено тестирование 322 новых образцов из 38 евразийских популяций. С учетом опубликованных данных сформирована выборка (полногеномные матрицы высокой плотности генотипирования), включающая 93 популяции, из которых 22 тюркоязычные.
Вопросы поставлены грамотно. На первый вопрос получен положительный ответ. Общий сигнал в генофонде тюрок имеется. Ответы на другие вопросы нам непонятны. Непонятны и выводы. «Finally, we used the ALDER method and inferred admixture dates (~9th–17th centuries) that overlap with the Turkic migrations of the 5th–16th centuries». То есть, те, кто принес сигнал (соответствующие генетические маркеры) в регионы западной Евразии, смешались с аборигенными популяциями в IX-XVII вв. Их потомки стали тюрками. Но это означает, что носители сигнала ранее IX в. из своей внутриазитской родины на запад не выходили, «migrations of the 5th-16th centuries» до IX в. не существовали. Рабочая гипотеза опровергнута. Однако, такие простые логические выводы авторы публикации не сделали. Вместе с тем они привели уникальные результаты грамотной обработки фактических данных. Представлялось целесообразным выполнить их независимую интерпретацию. Ее результаты изложены ниже.
2. Инструментарий популяционной генетики
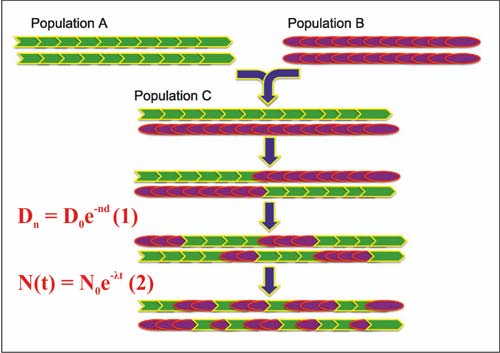
Рис. 1 – Иллюстрация процесса рекомбинации предковых геномов
при смешении двух популяций [Pugach et al., 2011, Fig. 1].
Формулы красным цветом вписаны А.М. Тюриным.
Пояснения по ним даны в тексте
При смешении двух популяций происходит процесс рекомбинации предковых геномов, по результатам которого они разбиваются на блоки (Рис. 1). Со временем блоки становятся все более узкими и равномерно распределяются вдоль и среди хромосом. То есть, количество и ширина блоков (аллель) является функцией времени, прошедшего с момента смешения, выраженного количеством поколений. Это дает возможность датировать событие «смешение двух популяций» только по генетическим данным.
Общая формула, являющаяся основой датирования события «смешение …», приведена на рисунке 1 (1). D0 – длина аллель в предковых популяциях; n – число поколений после смешения; Dn – длина аллель спустя n поколений после смешения; d – генетическая дистанция между предковыми популяциями [Loh et al., 2013]. Длительность поколения принимается равной 30 годам. Имеется две модификации метода – ALDER [Loh et al., 2013] и SPCO [Pugach et al., 2011]. У авторов публикации [Hellenthal et al., 2014] своя модификация. Мы просмотрели и другие публикации, в которых дано описание метода датирования, например, [Moorjani et al., 2011]. Поразительно, но в них в явном виде не фигурирует параметр «скорость процесса»! Предполагается, что процесс уменьшения длинны аллель от поколения к поколению подчиняется нормальному закону распределения вероятностей. Если это так, то он должен описываться формулой распада радиоактивных изотопов (Рис. 1, (2)), где t – время; N0 и N(t) – количество ядер в момент времени t = 0 и t; λ – вероятность распада ядра в единицу времени. Параметр d в формуле (1), просто обязан быть скоростью процесса. Но, просмотрев череду формул в отмеченных выше публикациях, нам не удалось разобраться в этом вопросе. Поэтому предположим, что скорость рекомбинации пропорциональна генетической дистанции между предковыми популяциями, оцененной специальным способом. Для нас важно, что генетики не имеют оценок скорости рассматриваемого процесса и «не знают», что только на их основе можно датировать событие «смешение …».
Вторая особенность модификаций датирования события «смешение …» – их тестирование на эталонных событиях (с достоверно известными датами) не выполнено. Исключением является датирование события «смешение негров с европейцами» на юго-западе США [Pugach et al., 2011]. В генофонде потомков негров-рабов 19 % европейских маркеров. По результатам датирования получена дата смешения – 6 поколений назад, с учетом доверительного интервала – 4-8 поколений. Она соответствует времени освоения этого региона плантаторами США. Метод работает. Но результаты тестирования ничего не говорят о возможных системных искажениях полученных дат. Кроме того, в этом случае мы имеем дело не с событием «смешение …», а с процессом, который продолжается и сегодня.
3. Генетический атлас событий «смешение популяций»
Здесь мы сделаем отступление от публикации [Yunusbayev et al., 2015]. Авторы статьи [Hellenthal et al., 2014] рассмотрели полногеномные маркеры выборки, включающей 95 популяций. В последние два тысячелетия выделили эпохи их смешения: европейская колонизация Америки (с 1492 г.), славянская и тюркская миграция (500-1100 гг.), арабская торговля рабами (650-1900 гг.), Монгольская (1206-1368 гг.) и Кхмерская (802-1431 гг.) империи. Славянская и тюркская миграция к рассматриваемому нами вопросу отношения не имеет. Ей затронуты греки, румыны, болгары, поляки, венгры, литовцы и белорусы. Миграция в эпоху Монгольской империи отразилась в генофондах тюркских популяций, включенных в выборку, – турки, узбеки и уйгуры. Отразилась она и в генофондах не тюркских популяций – лезгины, хазарейцы и монголы.
Монголы смешались в 1334 г. (1194-1446 гг.) с орочонами или якутами. Доля их маркеров составляет 36 %. Имеются и маркеры китайцев (хань). У хазарейцев доля маркеров монголов составляет 45 %. Смешение произошло в 1306 г. (1250-1390 гг.). Имеются маркеры иранцев. Смешение у предков турок произошло в 1250 г. (1166-1362 гг.), узбеков – 1362 г. (1306-1446 гг.), уйгуров – 1306 г. (1222-1390 гг.). В скобках указан доверительный интервал, для которого надёжность составляет 95 %. Доля монгольских маркеров в генофонде этих популяций составляет 8, 38 и 50 % соответственно. У турок имеются маркеры иранцев, а у узбеков и уйгуров – турок. Лезгины рассмотрены ниже.
Результаты нашей интерпретации рассмотренных данных сводятся к следующему.
1. Имеется ли в генофонде турок, узбеков и уйгуров общий сигнал? Да.
2. Связан ли он с доисторическими кочевыми тюрками? Нет. Он связан с монголами.
3. В генофондах трех популяций тюрок имеются свидетельства только одного события «смешение …», связанного с пришельцами с востока. Оно датировано второй половиной XII – первой половиной XV вв. Наиболее вероятная дата 1306 г.
4. Предполагаемые миграции кочевых тюрок в них не отражены.
5. В генофонде узбеков не отражены хорошо задокументированные переселения сообществ в период экспансии калмыков во второй четверти XVII – третьей четверти XVIII вв., а также экспансии джунгар в последней четверти XVII в. – 1758 г. Экспансия джунгар не отражена в генофонде уйгуров.
4. Генетическое наследие экспансии тюркоязычных кочевников
Исследования авторов публикации [Yunusbayev et al., 2015] включали четыре этапа:
- определение структуры полногеномных данных популяций, включенных в выборку, выделение у тюрок общего для них сигнала;
- выделение сигнала у тюрок на региональном фоне;
- датирование события «смешение двух популяций» в прошлом тюрок;
- интерпретация полученных результатов.
Структура полногеномных данных популяций определенна по технологии ADMIXTURE (Рис. 2). Выделено восемь кластеров. Цветом показана их доля в генофонде каждого индивидуума. Выборка разделена по регионам. На нижнем сегменте рисунка показаны колонки всей выборки, на верхнем – только тюркоязычных популяций. Они подписаны красным цветом. Например, в генофонде башкир суммарная доля полногеномные маркеров, попавших в кластеры К6 и К8, составляет примерно 22 %. «Родина» кластеров – Восточная Азия и Сибирь. В генофонде монголов Южной Сибири их доля составляет 78 %. Примерно 35 % в генофонде башкир составляет доля кластера К1. Его «родина» – Европа.
)
Рис. 2 – Структура полногеномных данных популяций Евразии выборки авторов публикации [Yunusbayev et al., 2015, Fig. 2]
Имеется ли в генофонде современных тюрок общий сигнал? Да, имеется. В генофондах тюрок западной Евразии он представлен кластерами К6 и К8, которые доминируют у тувинцев, бурятов, монголов и эвенков. Их генофонды приняты за эталоны при выделении сигнала на втором этапе исследований. Популяции сгруппированы по географическому принципу. В каждой из них показана доля маркеров эталонных популяций (Рис. 3). У гагаузов, уровень сигнала не превышает региональный фон. Достоверно выделяется сигнал у турок и азербайджанцев. Но его величина небольшая. Сигнал выделяется у кумыков и балкарцев. Но такая же его величина у не тюркоязычных адыгов и северных осетин. Четко выражен сигнал у кубанских ногайцев. У популяций Поволжья высокий региональный фон, соответствующий сигналу. Сигнал у татар и чувашей не выделяется на фоне не тюркоязычных коми, марийцев и удмуртов. У башкир сигнал проявился контрастно. У туркмен и таджиков сигнал имеет примерно одинаковую интенсивность. Отметим, что маркеры кластеров К6 и К8 у таджиков представлены вполне значимо. У узбеков, киргизов и казахов сигнал проявился суперконтрастно.
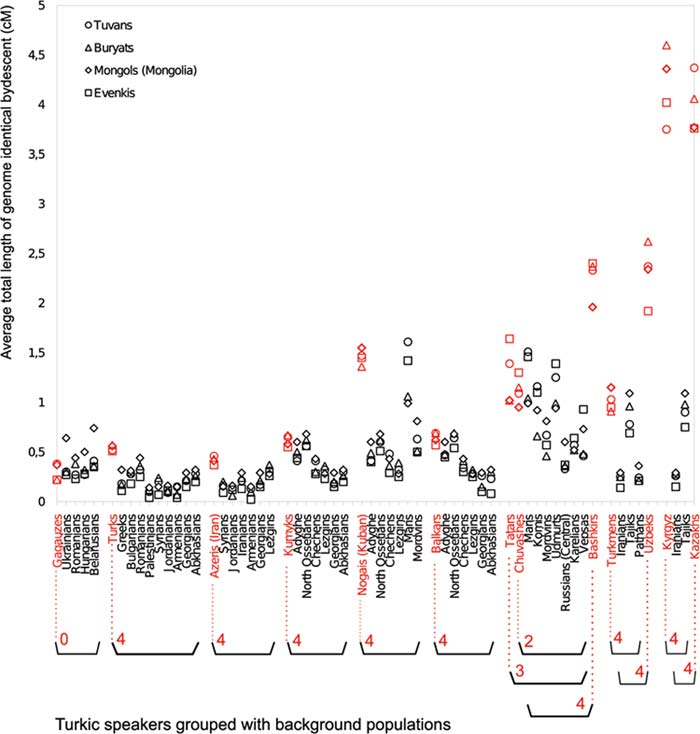
Рис. 3 – Доля генетических маркеров у популяций Евразии, идентичных с маркерами, доминирующими у тувинцев,
бурятов, монголов и эвенков [Yunusbayev et al., 2015, Fig. 4]
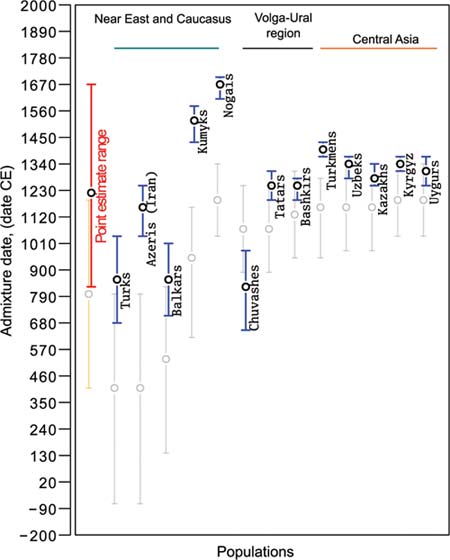
← Рис. 4 – Результаты датирования событий «смешение двух популяций» в прошлом тюрок [Yunusbayev et al., 2015, Fig. 5].
Доверительный интервал, для которого показаны погрешности, составляет 95 %.
Красный цвет – результаты ALDER, оранжевый – SPICO.
Даты до н. э. показаны со знаком «минус».
Итак, на втором этапе исследований определено, как именно сформировались современные тюркские народы. Это смешение местного субстрата, с теми, чей генофонд был близок тувинцам, бурятам, монголам и эвенкам. На третьем этапе выполнено датирование события «смешение двух популяций» (Рис. 4). Применено две модификации метода – ALDER и SPCO. Результаты совпали не в полной мере. Для их ранжирования выполнено моделирование процесса смешения. Даты, полученные по модификации ALDER, полностью соответствуют результатам моделирования. Даты, полученные по модификации SPCO, по отношению к ним системно смещены. По этой причине они исключены из дальнейшего рассмотрения. Выше отмечено, что результаты интерпретации нам непонятны. Непонятна и логика интерпретации фактических данных.
5. Авторская интерпретация результатов генетических исследований
5.1. Формальная интерпретация
В этом разделе приведена наша интерпретация данных, полученных авторами публикации [Yunusbayev et al., 2015]. Ими сделан вывод: «Finally, we used the ALDER method and inferred admixture dates (~9th–17th centuries)». Это не так. У татар, башкир, туркмен, узбеков, казахов, киргизов и уйгуров событие «смешение …» произошло в 1185-1435 гг. (Рис. 4). С учетом доверительного интервала к этому же интервалу можно отнести и рассматриваемое событие у азербайджанцев. Наиболее вероятная дата смешения – 1300 г. То есть, в генофонде подавляющего числа тюркоязычных популяций не имеется сигнала гипотетических кочевых тюрок V-XII вв., генофонд которых близок тувинцам, бурятам, монголам и эвенкам. Не имеется их сигнала у кумыков и ногайцев.
Даты событий «смешение …» у турок, чувашей и балкарцев попали в более ранний интервал времени, чем у большинства тюркоязычных популяций. У турок это событие датировано 873 (692-1054 гг.). Кочевые тюрки могли пройти в Малую Азию либо через Среднюю Азию, либо через Восточную Европу. Но у тюркоязычных популяций в этих регионах соответствующего сигнала не имеется. Это указывает на то, что дата авторов публикации [Yunusbayev et al., 2015] не верна. Условно верная дата – 1250 г. (1166-1362 гг.), у авторов публикации [Hellenthal et al., 2014]. Это тот же интервал, в который попали аналогичные события у подавляющего числа популяций тюрков. У чувашей событие «смешение …» датировано 816 г. (677-955 гг.). Авторы последней публикации для чувашей получили три разные даты рассматриваемого события – 1606 г. до н. э., 886 г. (690-1054 гг.) и 1502 г. (1418-1670 гг.). Смешение их предков произошло с якутами или монголами. То есть, методы датирования события у предков чувашей дают неустойчивые результаты. Ни одна из дат не может быть принята во внимание. Про достоверность даты этого события у предков балкарцев – 864 г. (727-1003 гг.), сказать ничего не можем. Тем не менее, при обобщении результатов датирования события «смешение …» ее следует идентифицировать как «выброс», то есть, как недостоверную.
Первый наш вывод определен. В генофонде современных тюрок имеется общий сигнал. По технологии ALDER он датирован периодом не ранее конца XII в. Это исключает его отнесение к доисторическим кочевым тюркам. Отметим, что вывод сделан только путем формальной интерпретации результатов генетических исследований, полученных авторами публикации [Yunusbayev et al., 2015].
Генофонды тувинцев, бурятов, монголов и эвенков приняты за эталоны при выделении сигнала на втором этапе исследований. Но генофонд эвенков (Рис. 2) не соответствует общему сигналу у тюрок. В генофондах почти всех тюрок, кроме гагаузов, долган и якутов, имеются полногеномные маркеры кластера К8. А у эвенков их нет. Генофонды бурятов и монголов Сибири неразличимы. Но в генофонде монголов Монголии минимальное число маркеров, не относящихся к кластерам К6 и К8. Генофонд тувинцев отличается от них только наличием маркеров кластера К5. Два наших вывода определенные.
1. Общий генетический сигнал у тюрков кроме гагаузов, долган и якутов связан с монголами. Это ровно то, к чему пришли авторы публикации [Hellenthal et al., 2014]. В прошлом турок, узбеков и уйгуров было смешение их предков с монголами.
2. Сигнал «смешение с монголами» имеется и у не тюркоговорящих популяций – коми, марийцев, удмуртов, адыгов, северных осетин и таджиков.
Понять, как работают эталоны можно на примере популяций Поволжья. У татар и марийцев величина сигналов «эвенки» и «тувинцы» доминирует над величиной сигналов «монголы» и «буряты» (Рис. 3). Связано это с наличием в генофонде татар, марийцев, эвенков и тувинцев маркеров кластера К5 (Рис. 2), который, в первом приближении, можно идентифицировать с уралоидностью. А величина сигналов «монголы» и «буряты» у татар, чувашей, марийцев и удмуртов практически одинаковая. То есть, две тюркоязычные популяции не выделяются на фоне своих соседей.
С долганами и якутами все понятно. Они обособились в северных районах Евразии до того, как сформировался поток «китайских» генетических маркеров в Центральную Азию. Это маркеры кластера К8. Монголы начали мигрировать на запад только после того, как получили их в свой генофонд.
5.2. Датирование событий «смешение двух популяций»
5.2.1. Постановка задачи
Напомним, что две технологии датирования события «смешение …» – ALDER и SPICO, не прошли соответствующее тестирование на реальных генетических данных. Даты событий «смешение …», полученные по модификации ALDER, полностью совпали с результатами моделирования. Это свидетельствует только о том, что датирование и моделирование выполнены по одному и тому же алгоритму, и вовсе не означает соответствие полученных дат реалиям. Исходя из этого, задача проста и понятна. Нужно датировать рассматриваемое событие в прошлом тюркоязычных популяций другими методами. Сделать это можно по результатам идентификации общего сигнала в их генофондах. Он связан с монголами.
Авторы публикации [Yunusbayev et al., 2015] рассматривают только часть генетического сигнала. Генофонд популяций включает частоты гаплогрупп Y-хромосомы (передаются по мужской линии), мт-ДНК (передаются по женской линии), аутосомные и полногеномные маркеры (передаются и по мужской, и по женской линиям). В соответствии с Традиционной историей, миграции с востока в зону проживания тюркоязычных популяций в западных регионах Евразии осуществили монголы в середине XIII в. и калмыки в первой половине XVII в. А калмыки – это ойраты, западные монголы. То есть, генофонды монголов XIII в. и калмыков XVII в. практически неразличимы. Доминирующая по численности этническая группа монголов – халхи. В характеризующей их выборке (N=85) частоты гаплогруппы С – 56,5 %, О3 – 18,8 %, D – 3,5 % [Katoh et al., 2005]. У северных китайцев (N=42) частоты этих гаплогрупп 9,5, 52,4 и 0 % соответственно. Гаплогруппа D доминирует у японцев (N=117) – 38,3 %. Понятно, что гаплогруппа О3 попала к монголам от китайцев, а D – от японцев. Эти три гаплогруппы в западной Евразии являются генетическими маркерами монголов и калмыков. Их же маркерами являются восточноевразийские гаплогруппы мт-ДНК. То есть, в общем случае инструментарием популяционной генетики нам не удастся идентифицировать события «смешение …» в прошлом тюркоязычных популяций. Они могли смешаться с монголами и (или) с калмыками. Но имеются другие методы датирования этих событий.
5.2.2. Датирование по литовским татарам
Литовские татары, как субэтнос, сформировались из военного сословия политических образований XV – начала XVII в. – Большой Ногайской орды, Малой Ногайской орды и Крымского ханства, являющихся осколками Улуса Джучи XIII-XIV вв. Обособились в Великом княжестве Литовском в начале XVI в. У литовских татар (N=74 и N=35) не выявлены «монгольские» гаплогруппы С, О и D. Наличие монголов по мужской линии в их предках исключается [Тюрин, 2017-а]. Такое могло быть только в одном случае – монголы не приходили в Восточную Европу и сопредельные регионы Азии ранее XVI в.
5.2.3. Датирование по казахским торе
В соответствии с родословными и семейными преданиями казахские торе (потомки правителей региона) считают Чингисхана свои прямым предком. Среди тестированных торе (N=23) отмечены носители разных гаплогрупп Y-хромосомы [Адамов, Сабитов, 2015]. Они не являются потомками одного родителя, который жил в последние тысячелетия. Среди торе 8 носителей гаплогруппы C. Их гаплотипы формируют кластер. Датирование кластера генохронологическим методом дает время жизни его прародителя 225-270 лет назад [Тюрин, 2017-б]. То есть, прародитель или его потомки влились в клан торе не ранее XVIII в. Основатель кластера гаплотипов гаплогруппы C был калмыком или джунгаром. Таким образом, и у казахских торе до прихода в регион калмыков не имелось носителей гаплогруппы С. По особенностям их генофонда общий сигнал у тюрок идентифицируется однозначно. Он связан с калмыками.
5.2.4. Датирование по якутам
Выше отмечено, что якуты обособились в регионе их сегодняшнего проживания до того, как сформировался поток «китайских» генетических маркеров в Центральную Азию. Это относится и к гаплогруппе О. Ее гаплотипы (N=659) у якутов не выявлены [Адамов, 2010]. Взаимосвязанные процессы – первый этап этногенеза якутов и начало колонизации территории Якутии, датированы нами по историческим, археологическим и генетическим (гаплотипы Y-хромосомы) данным, а также по динамике изменения численности популяций [Тюрин, 2010-а]. Первый этап этногенеза якутов – XVI в. Начало колонизации территории Якутии – вторая половина XVI в. То есть, в первой половине этого века у популяций северных регионов Центральной Азии, родины якутов, включая их ближайших генетических «родственников» монголов, еще не имелось «китайских» генетических маркеров. А у монголов, пришедших в Восточную Европу, они имелись. Следовательно, монголы начали движение на запад не ранее XVI в.
5.2.5. Датирование по хазарейцам
По результатам изучения гаплотипов гаплогруппы С Y-хромосомы популяций Средней Азии, Внутренней и внешней Монголии выделен кластер, который назван Star Cluster [Zerjal et al., 2003]. Соответствующие ему гаплотипы имеются примерно у 16 миллионов мужчин региона, что составляет 8 % его мужского населения, или 0,5 % населения мира. Генохронологическое датирование дало годы жизни прародителя кластера, совпадающие с годами жизни Чингисхана. Сделано предположение, что 16 миллионов мужчин региона являются его прямыми потомками. В последующих публикациях Star Cluster называют и кластер Genghis Khan. Мы выполнили датирование прародителя кластера двумя способами [Тюрин, 2010-б]. Генохронологическое датирование дает вторую половину XV – XVI вв. Датирование по историческим данным (по калмыкам, у которых выявлен всего один гаплотип кластера Genghis Khan) – не ранее рубежа XVI и XVII вв. Исходя из этого, кластер Genghis Khan не может быть отнесен к Чингисхану. Логика нашего датирования проста и понятна. Предки калмыков вышли из Западной Монголии в самом начале XVII в. Если у сегодняшних калмыков практически не имеются гаплотипы кластера Genghis Khan, то их в то время не имелось и в Монголии. Один выявленный у калмыков носитель этого гаплотипа, скорее всего, является потомком зюнгаров, которые пришли в регион их проживания в Прикаспии после разгрома китайцами Джунгарского ханства в 1755-1759 гг.
Наибольшее число носителей гаплотипов кластера Genghis Khan (порядка 40 %) выявлено у хазарейцев. При учете нашей даты его прародителя (по калмыкам), предки хазарейцев не могли выйти с территории Монголии ранее XVII вв. Выявленное у них событие «смешение …» произошло не ранее этой даты. Напомним, что оно датировано авторами публикации [Hellenthal et al., 2014] 1306 г. (1250-1390 гг.). То есть, методы популяционной генетики датирования событий «смешение …» дают систематическую погрешность. В данном случае, дата, полученная на их основе, древней реальной не менее, чем на 350 лет.
Скорее всего, хазарейцы – это потомки части джунгар, перешедших через Бадахшан на территорию Афганистана после разгрома Джунгарского ханства. Мы посещали и китайскую часть Памира со стороны Кашгара, и таджикскую со стороны Душанбе. Путь из Кашгара в Афганистан через Памир не только вполне проходим для кочевников, но и удобен. Не имеется недостатки ни в корме для их стад, ни в воде. Религией Джунгарского ханства был буддизм. Исходя из этого, понятно, кто и когда построил знаменитые статуи Будды в афганской провинции Бамиан, одном из центров компактного проживания хазарейцев – их предки джунгары во второй половине XVIII в., до того, как приняли ислам.
5.2.6. Датирование по историческим и археологическим данным
Автор монографии [Трепавлов, 2016, с. 474] дал справку по письменным свидетельствам: «ни малейшего следа присутствия монголов среди предков ногаев не зафиксировано какими-либо источниками». Имеются в виду ногаи Большой ногайской орды по ее состоянию на начало XVII в.
По результатам рассмотрения данных по 276 погребениям кочевников XIII-XIV вв. степного Приуралья (рассматриваемый регион включает и Южное Зауралье) сделано заключение: «вопрос о наличии в регионе погребений самих монголов остается открытым» [Матюшко, 2011, с. 286]. То есть, погребений монголов в регионах, сопредельных с ареалом расселения башкир, не выявлено. Погребениям кочевников Приуралья XIII-XIV вв. соответствует поздний этап чияликской археологической культуры Южного Урала. Мы просмотрели несколько публикаций ([Гарустович, 2015] и др.). Какого-либо влияния на чияликскую культуру монголов не отмечается.
5.2.7. Результаты датирования
Установлено, что события «смешения …» в прошлом тюркоязычных популяций западной Евразии связаны с монголами. Инструментарием популяционной генетики они датированы 1185-1435 гг. По результатам датирования этих событий шестью независимыми методами получено, что они не могут быть отнесены к монгольской экспансии времен Монгольской империи (1206-1368 гг.). Они связаны с экспансией калмыков XVII – третьей четверти XVIII вв. Даты, полученные методом генетиков, имеют систематические погрешности.
6. Особенности генофонда некоторых популяций
6.1. Лезгины
В 1306 г. (1138-1446 гг.) произошло смешение предков лезгин с уйгурами, а по другому варианту обработки данных – с индийскими евреями [Hellenthal et al., 2014]. Доля их генетических маркеров у современных лезгин 10 %. Но было еще одно смешение – с турками или шотландцами. Его параметры не определены. Ничего необычного в таком раскладе мы не видим. При существенном расширении выборки, уйгуров могут сменить, например, караногайцы, а индийских евреев – горские евреи. С турками проблем не имеется. Но шотландцы «ведут себя странно». По одному из вариантов обработки данных, у калашей 35 % их маркеров. Смешение произошло в 738 г. (1326-94 гг.) или 598 г. (990-206 гг.) до н. э. Смешение лезгин с «уйгурами» произошло не ранее прихода калмыков в северо-западный Прикаспий (середина XVII в.).
6.2. Башкиры
У башкир в пяти выборках (N=935) выявлено 28 носителей «монгольских» гаплогрупп Y-хромосомы С и О [Тюрин, 2018-а, 2019], что составляет 3,0 %. В среднем у башкир 39,2 % восточноевразийских гаплогрупп мтДНК [Лобов, 2009]. По этим данным можно ожидать, что у них должно быть 21 % «монгольских (калмыцких)» полногеномных маркеров. А их примерно 22 % (Рис. 2, кластеры К6 и К8). У башкир полное совпадение двух разных независимых массивов генетических данных. Авторы публикации [Балановская и др., 2017] установили, что в их генофонде имеется еще один «след» миграции из Восточной Сибири. Это ветвь N3a2 гаплогруппы Y-хромосомы N3. По частотам она доминирует у клана еней (44 %), но отсутствует у остальных шести кланов северо-западных башкир. Скорее всего, ветви N3a2 соответствует кластер полногеномных маркеров К5, доминирующий по частотам у ненцев, манси, кетов и нганасан. Частоты этого кластера у башкир порядка 10 %.
По результатам анализа естественнонаучных данных (краниология и популяционная генетика), исторических свидетельств, родовых и клановых этнонимов установлено, что башкиры являются европеоидами с небольшой долей центрально-азиатской монголоидности, которую они получили, в основном, через невест казашек и калмычек. В меньшей мере – за счет интеграции в свои кланы ногаев, метисированных при контактах с калмыками, и калмыков [Тюрин, 2018-а]. Доля центрально-азиатской монголоидности у башкир составляет 21-22 %. Этот параметр оценен по двум независимым массивам генетических маркеров
6.3. Казахи
В генофонде казахов доля маркеров кластеров К6 и К8 составляет примерно 60 %. Младший жуз состоит из трех родовых объединений – байулы, алимулы и жетыру. Два первых объединения формируют союз алшин (алчин). В публикации [Баймуханов, Баимбетов, 2018] приведены результаты ДНК-тестирования казахов объединения алимулы (N=51), которым охвачены все его шесть родов (шомекей, шекты, каракесек, торткара, карасакал и кете). Частота пяти линий гаплогруппы С составляет 92,2 %, в том числе линии C-Y15552 – 76,5 %. Оставшиеся четыре образца дали гаплогруппы R1 и G. Линия C-Y15552 имеет высокие частоты и у объединения байулы. Всего у алшинов (N=189) 80,5 % носителей гаплогруппы С [Сабитов, Жабагин, 2015]. Датирование выявленной у казахов алимулы специфической мутации в Y-хромосоме по технологии TMRCA дало время ее появления – «550 лет назад» [Баймуханов, Баимбетов, 2018]. В публикации приведены и гаплотипы тестированных. Гаплотипы линии C-Y15552 у алимулы формируют кластер (его называют алчинским). Наше датирование его основателя по числу мутаций относительно базового гаплотипа дало «640 лет назад».
Мы видим только один вариант этнической интерпретации полученных данных. Погрешность двух независимых способов датирования составляет примерно +/- 100 лет. Примем 1980 г. за среднюю дату рождения тестированных. Основатель кластера гаплотипов с характерной мутацией в Y-хромосоме родился на территории Западной Монголии в период со второй половины XIII по первую половину XVI вв. Был он ойратом. В начале XVII в. его потомки пошли на север, а потом на запад. Их стали называть калмыками. В 20-30-х годах они заняли территорию (кроме Мангышлака), на которой сегодня проживают казахи объединений алимулы и байулы. В конце XVII – начале XVIII вв. калмыки, потомки основателя алчинского кластера стали казахами.
В 1731 г. хан Абулхаир от имени Младшего жуза принял российское подданство. До этой даты о казахах жуза имеются только отрывочные свидетельства. Первое упоминание объединения алимулы относится к 1748 г., но названо оно каракесек [Баймуханов, Баимбетов, 2018]. Что это именно то объединение, которое стало называться алимулы, говорят названия его пяти родов. Они совпадают с названиями пяти родов алимулы. В столетний период между двумя датами – 20-е годы XVII в. и 1731 гг., в Южном Зауралье, северном Приаралье и северо-восточном Прикаспии (к востоку от Урала) происходили какие-то этнические процессы, о которые почти ничего не известно. По их результатам были сформированы родовые объединения казахов алимулы и байулы.
6.4. Ногайцы
В публикации [Тюрин, 2017-в] рассмотрены многочисленные переселения ногаев Большой ногайской орды, рода которых вошли в калмыцкие улусы и перешли с ними Волгу. От калмыков они получили монгольские гаплогруппы и центрально-азиатский тип монголоидности [Тюрин, 2018-б]. У кубанских ногайцев (N=90) по частотам доминирует гаплогруппа R1a1a-M198 – 48 % [Схаляхо, Чухряева, 2016]. Частоты ее «славянской» субветви R1a1a1g-M458 – 13 %. Частоты монгольских гаплогрупп – 4 %, в том числе: С – 3 % и О – 1 %. По другой выборке [Литвинов, 2010] у кубанских ногайцев (N=87) частота монгольских гаплогрупп – 12,8 %, в том числе: С – 8,1 %, О – 3,5 %, D – 1,2 %. По двум выборкам частота монгольских гаплогрупп у ногайцев составляет 8,5 %, в том числе гаплогруппа С – 5,6 %. Обращаем внимание на существенно отличные результаты двух выборок в части частот монгольских гаплогрупп. По полногеномным данным у ногайцев примерно 20 % монгольских маркеров (Рис. 2). Скорее всего, у них существовали брачные отношения с калмыками. То есть, количество монгольских маркеров, попавших к ним по женской линии, выше, чем по мужской.
В публикациях высказывается мнение, что часть кубанских ногайцев являются потомками джетисанцев и джембуйлуков Большой ногайской орды. Они вошли в калмыцкие улусы и вместе с ними перешли Волгу. Но в выборке ногайцев из публикации [Yunusbayev at al, 2015] доминируют полногеномные маркеры кавказского кластера К3 (Рис. 2). Последний факт не в полной мере соответствует и доминированию у них гаплогруппы R1a. Для того, чтобы разобраться в этом нужно знать, как сформированы рассмотренные выборки. То есть, вопросе нуждается в специальном рассмотрении.
Можно предположить, что джетисанцы и джембуйлуки Большой ногайской орды до вхождения в нее проживали на Северном Кавказе. Примерно во второй четверти XVII в. их рода были включены в калмыцкие улусы. В 20-х годах XVIII в. они вышли из подчинения калмыкам и были переведены Крымским ханством в Белгородскую орду [Тюрин, 2017-в]. Таким образом, в плотном контакте с калмыками ногаи пробыли примерно 90 лет, что составляет три поколения. Это уже не событие «смешение двух популяций», а процесс. Скорее всего, по этой причине дата события «смешение …», определенная по технологии ALDER (Рис. 4), существенно отличается от дат этого события в других тюркоязычных популяциях.
6.5. Турки
В публикации [Cinnioglu et al., 2004] приведены результаты тестирования турок (N=523, 9 регионов). Всего выявлено 7 носителей гаплогруппы С и 1 – О, что составляет 1,5 %. Причем, 4 (4,9 %) носителя гаплогруппы C живут в Стамбуле (N=81). В XVIII-XIX вв. в Турцию происходила массовая миграция ногаев, причем, именно тех, чьи предки десятилетия провели в калмыцких улусах [Тюрин, 2017-в, 2020]. Их миграция осуществлялась напрямую с Кубани, а также через Крым и Добруджу. Но относительно высокие частоты носителей монгольских гаплогрупп в Турции невозможно объяснить только миграцией туда ногаев, метесированных при контактах с калмыками. Скорее всего, существовал и поток калмыков (ясырь) с Северного Кавказа и Крыма.
7. Общие выводы
В пункте 1 приведено состояние инструментария популяционной генетики применительно к рассматриваемому вопросу, в пункте 2 – результаты исследований авторов публикаций [Hellenthal et al., 2014; Yunusbayev et al., 2015], в пунктах 3-7 – результаты интерпретации данных популяционной генетики, полученные автором настоящей статьи.
1. В популяционной генетике создан инструментарий, позволяющий выполнять анализ массивов полногеномных маркеров, характеризующих выборки популяций, выделять события «смешение двух популяций» и их датировать. Однако тестирование способов датирования на эталонных событиях (с достоверно известными датами) пока не выполнено.
2. В генофондах тюркоязычных популяций имеется общий сигнал (совокупность генетических маркеров). По одной из версии его интерпретации у турок, узбеков и уйгуров он связан с монголами [Hellenthal et al., 2014]. Событие «смешение …» датировано второй половиной XII – первой половиной XV вв. Наиболее вероятная дата – 1306 г. По другой версии (рассмотрены данные по 22 тюркоязычным популяциям), сигнал сформирован на территории Монголии, Южной Сибири и Северного Китая. Соответствующие ему маркеры доминирует у тувинцев, бурятов, монголов и эвенков [Yunusbayev et al., 2015]. Событие «смешение …» датировано IX-XVII вв. Но в соответствии с фактическими результатами, это событие у предков татар, башкир, туркмен, узбеков, казахов, киргизов, уйгуров и азербайджанцев произошло в 1185-1435 гг. Наиболее вероятная дата – 1300 г.
3. У тюркоязычных популяций западной Евразии выявлено только одно событие «смешение …» с пришельцами с востока. Они однозначно идентифицированы как монголы. Но в соответствии с Традиционной историей, их миграции было две – монголы в середине XIII в. и калмыки в первой половине XVII в. Их генофонды неразличимы.
4. Авторское датирование события «смешение …» выполнено шестью независимыми способами: по особенностям генофонда литовских татар, казахских торе, якутов и хазарейцев, а также по историческим и археологическим данным. Приход монголов в западные регионы Евразии ранее XVI в. исключается. Исходя из этого, идентификация сигнала у тюрков однозначная. Он связан с калмыками.
5. В генофонде тюрок запада Евразии не имеется других сигналов, которые можно было бы связать с пришельцами с территории Монголии, Южной Сибири и Северного Китая. Логический вывод однозначен. Первой популяцией, которая дошла из этих регионов до западной Евразии, были калмыки. Этим опровергнута одна из составных частей Традиционной истории, называемой «Миграции кочевых тюрков V-XVI вв.» Калмыки принесли в западные регионы Евразии центрально-азиатский тип монголоидности, а также монгольские генетические маркеры – гаплогруппы Y-хромосомы С, О и D, восточноевразийские гаплогруппы мт-ДНК и полногеномные маркеры.
6. За генетическое наследие экспансии тюркоязычных кочевников авторы публикаций [Yunusbayev et al., 2015] приняли калмыцкий «след» у популяций (не только тюркоязычных) западных регионов Евразии.
7. Вывод по пункту 5 получен по массивам данных популяционной генетики. Он разрушает основы всех известных нам гипотез возникновения и распространения тюркского языка, кроме одной. При создании огромной Империи XIII-XVII вв. «На местные наречия колонизированных территорий наложились русский и тюркский языки ордынско-атаманских завоевателей. Оба языка долгое время были в ходу на всей территории Великой Империи и сформировали исходную языковую картину в целом» [Носовский и др., 2012]. Для многих популяций Евразии тюркский язык стал родным.
Литература
Адамов Д. Заметки о гаплогруппах Y-хромосомы якутов // Тhе Russian Journal of Genetic Genealogy, 2010, Vol. 2, № 1, с. 3-14.
Адамов Д. Сабитов Ж.М. Расчет TMRCA казахских родов // Тhе Russian Journal of Genetic Genealogy, Volume 7, 2015, № 2, с.14-19.
Баймуханов Н., Баимбетов Г. Генетический субклад ZQ5 гаплогруппы C-Y15552 родоплеменного объединения «Алимулы» // Journal of history, 2018, №2 (89), с. 172-180.
Балановская Е.В., Юсупов Ю.М., Схаляхо Р.А., Степанов Г.Д., Асылгужин Р.Р., Жабагин М.К., Балаганская О.А, Султанова Г.Д., Борисова Е.Б., Дараган Д.М., Балановский О.П. Генетические портреты семи кланов северо-западных башкир: вклад финно-угорского компонента в генофонд башкир // Вестник Московского университета. Серия XXIII. Антропология, 2017, № 3, с. 94-103.
Гарустович Г.Н. Чияликская археологическая культура эпохи средневековья на Южном Урале // Уфимский археологический вестник, 2015, № 15, с. 181-198.
Литвинов С.С. Изучение генетической структуры народов Западного Кавказа по данным о полиморфизме Y-хромосомы, митохондриальной ДНК и alu-инсерций. Автореферат диссертации на соискание ученой степени кандидата биологических наук. Уфа, 2010, 23 с.
Лобов А.С. Структура генофонда субпопуляций башкир. Автореферат диссертации на соискание ученой степени кандидата биологических наук. Уфа, 2009, 23 с.
Матюшко И.В. Особенности погребального обряда кочевников степного Приуралья XIII-XIV вв. // Известия Самарского научного центраРоссийской академии наук, 2011, Т. 13, № 3-1, с. 280-283.
Носовский Г.В., Фоменко А.Т., Фоменко. Т.Н. Русские корни «древней» латыни. Языки и письменность Великой Империи, 2012, Астрель, 606 с.
Сабитов Ж.М., Жабагин М.К. Этногенез казахов с точки зрения популяционной генетики. Вопросы конституционного строительства и роль лидера нации // III конгресс историков Казахстана, 2015, с. 375-379.
Схаляхо Р.А., Чухряева М.И. Генофонды ногайцев в контексте населения степного пояса Евразии (по маркерам Y-хромосомы) // Золотоордынская цивилизация, 2016, № 9, с. 326-333.
Трепавлов, В.В. История Ногайской Орды. 2-е изд., испр. и доп. Казань: Издательский дом «Казанская недвижимость», 2016, 764 с.
Тюрин А.М. Датирование первого этапа этногенеза якутов и начала колонизации территории Якутии // Электронный сборник статей «Новая Хронология», 2010-а, Вып. 10. [Новая хронология]
Тюрин А.М. Датирование кластера гаплотипов Y-хромосомы «Genghis Khan» // Электронный сборник статей «Новая Хронология», 2010-б, Вып. 10. [Новая хронология]
Тюрин А.М. Генетический портрет литовских татар и феномен «Монгольские завоевания 13 века» // Вестник Оренбургского государственного университета, 2017-а, № 5, с. 78-82.
Тюрин А.М. Казахские торе не являются потомками Чингисхана // Цивилизация знаний: российские реалии: труды Восемнадцатой Международной научной конференции, Москва, 21-22 апреля 2017 г. М.: РосНОУ, 2017-б, с. 93-94.
Тюрин А.М. Калмыки, караногайцы, кубанские ногайцы и крымские татары – геногеографический и геногенеалогический аспекты // Журнал фронтирных исследований, 2017-в, № 2, с. 7-29.
Тюрин А.М. К вопросу о монголоидных компонентах у башкир // Электронный сборник статей «Новая Хронология», 2018-а, Вып. 15. [Новая хронология]
Тюрин А.М. Ногаи: антропологический аспект // Электронный сборник статей «Новая Хронология», 2018-б, Вып. 15. [Новая хронология]
Тюрин А.М. Ямная археологическая культура Южного Урала и Волго-Уральского региона (вторая четверть XVII – XVIII вв. н. э.) создана башкирами-бурзянами // Электронный сборник статей «Новая Хронология», 2020, Вып. 17. [Новая хронология]
Cinnioglu C., King R., Kivisild T., Kalfoglu E., Atasoy S., Cavalleri G.L., Lillie A.S., Roseman C.C., Lin A.A., Prince K., Oefner P.J., Shen P., Semino O., Cavalli-Sforza L.L., Underhill P.A. Excavating Y-chromosome haplotype strata in Anatolia. Hum Genet. 2004 Jan;114(2):127-48. Epub 2003 Oct 29.
Hellenthal G., Busby G.B., Band G., Wilson J.F., Capelli C., Falush D., et al. A genetic atlas of human admixture history. Science. 2014;.343(6172):747–51. pmid:24531965
Katoh T., Munkhbat B., Tounai K., Mano S., Ando H., Oyungerel G., Chae G.-T., Han H., Jia G.-J., Tokunaga K., Munkhtuvshin N., Tamiya G. and Inoko H. Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis. Gene, Volume 346, 14 February 2005, pp. 63-70.
Loh P.R., Lipson M., Patterson N., Moorjani P., Pickrell JюK., Reich D., et al. Inferring admixture histories of human populations using linkage disequilibrium. Genetics. 2013;193(4):1233–54. pmid:23410830
Moorjani P., Patterson N., Hirschhorn J., Keinan A., Hao L., et al., 2011. The history of African gene flow into Southern Europeans, Levantines, and Jews. PLoS Genet. 7(4):e1001373
Pugach I., Matveyev R., Wollstein A., Kayser M., Stoneking M. Dating the age of admixture via wavelet transform analysis of genome-wide data. Genome Biology. 2011;12(2):R19. pmid:21352535
Yunusbayev B., Metspalu M., Metspalu E., Valeev A., Litvinov S., Valiev R., Akhmetova V., Balanovska E., Balanovsky O., Turdikulova S., Dalimova D., Nymadawa P., Bahmanimehr A., Sahakyan H., Tambets K., Fedorova S., Barashkov N., Khidiyatova I., Mihailov E., Khusainova R., Damba L., Derenko M., Malyarchuk B., Osipova L., Voevoda M., Yepiskoposyan L., Kivisild T., Khusnutdinova E., Villems R. The genetic legacy of the expansion of Turkic-speaking nomads across Eurasia. // PLoS Genet. – 2015. V.11(4):e1005068. Impact factor (Thomson Reuters) - 7,528. doi: 10.1371/journal.pgen.1005068
Zerjal T., Xue Y., Bertorelle G., Wells R.S., Bao W., Zhu S., Qamar R., Ayub Q., Mohyuddin A., Fu S., Li P., Yuldasheva N., Ruzibakiev R., Xu J., Shu Q., Du R., Yang H., Hurles M.E., Robinson E., Gerelsaikhan T., Dashnyam B., Mehdi S.Q., Tyler-Smith C. The genetic legacy of the Mongols. Am J Hum Genet. 2003 Mar;72(3):717-21. Epub 2003 Jan 17.
статья получена 27.01.2019
