УДК 575.17+572.2
Венгры, мадьяры и авары: популяционная генетика, антропология, этнонимы, лингвистика, этнические реконструкции
А.М. Тюрин
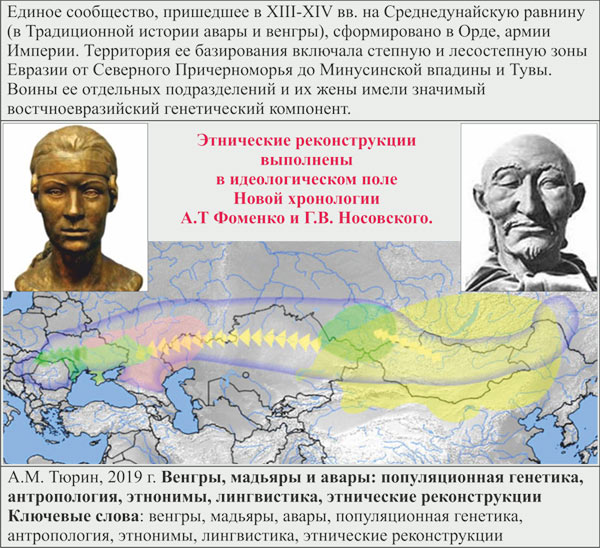
Ключевые слова: венгры, мадьяры, авары, популяционная генетика, антропология, этнонимы, лингвистика, этнические реконструкции.
1. Постановка задачи
В соответствии с Традиционной историей предками венгров являются автохтонное население Среднедунайской равнины и пришельцы с востока – скифы, сарматы, гунны, авары, оногуры, булгары, мадьяры, а также другие поздние кочевники. Венгерский язык относится к финно-угорской группе уральской языковой семьи. Ранее мы пытались выделить этнические компоненты у венгров по данным популяционной генетики по состоянию на 2010 г. [Тюрин, 2000, Венгры]. Наши выводы: «венгры не имеют значимых генетических отличий от народов Центральной Европы и Балкан. Однако среди них имеется небольшое число носителей гаплогрупп Y-хромосомы и мтДНК (порядка 1%), характерных для сибирских и центральноазиатских народов, а также гаплотипов западноевразийских гаплогрупп, совпадающих с гаплотипами останков из курганных захоронений Южной Сибири». В последние два года опубликованы статьи, в которых приведены генофонды венгров, мадьяр и аваров [Neparaczki, 2018; Neparaczki, 2019; Post, 2019]. Представляется целесообразным выполнить их независимый анализ и соответствующие этнические реконструкции. Нами рассмотрены данные популяционной генетики, краниологии, этнонимы сообществ, пришедших на Среднедунайскую равнину,и некоторые особенности венгерского языка. Данные популяционной генетики включают гаплогруппы Y-хромосомы (передаются по мужской линии), мтДНК (передаются по женской линии), а также аутосомные и полногеномные маркеры (передаются по мужской и женской линиям). В статьях на английском языке применяется термин «Hungarian Conquerors». Мы будем пользоваться российским термином «мадьяры».
Гаплогруппы C, O и D мы называем «монгольскими». У монголов (N=852) их частоты 58,1, 16,7 и 0,8 % соответственно [Дамба, 2018], у калмыков (N=207) частоты первых двух гаплогрупп 59,4 и 4,5 % [Balinova, 2019]. Они являются генетическими маркерами (от термина «генезис») монголов (калмыков) в Восточной Европе и сопредельных районах Азии. Они же являются генетическими маркерами центрально-азиатской монголоидности в регионах к западу от Центральной Азии. Гаплогруппа С является генетическим маркером собственно монголов. Гаплогруппа О по частоте доминирует у китайцев, гаплогруппа D – у корейцев и японцев. Эти две гаплогруппы попали к монголам от соседних популяций.
У всех гаплогрупп Y-хромосомы выделены субклады. В гаплогруппах мтДНК выделяются западно- и восточноевразийские. Аутосомные и полногеномные маркеры популяций в поле главных компонент определяют их этническое взаимоотношение. По этим данным (при комплексном их рассмотрении) можно идентифицировать расовую принадлежность популяций, сообществ и отдельных индивидов, выделять у них конкретные этнические компоненты. Первые результаты такого подхода изложены в наших публикациях [Тюрин, 2019, Караногайцы; 2019, Кубанские ногайцы 2019; Викинги, 2019; Скифы, 2019, Башкиры-1, 2019; Башкиры-2, 2019].
По гаплогруппам Y-хромосомы и мтДНК венгры не отличаются от соседних с ними популяций. Это научный факт. Дополнительно касаться этого вопроса мы не будем. Рассмотрим только особенности их генофонда.
2. Этнические группы венгров
Гунны пришли в Восточную Европу с востока. Их владения при Аттиле (434-453 гг.) простирались от Волги и Кавказа до Рейна. Включали и Среднедунайскую равнину. Авары (в русских свидетельствах обры) пришли на Среднедунайскую равнину тоже с востока. В 562-568 гг. они подчинили местное население – славян, германцев и романизированные сообщества. Сформировался Аварский каганат, просуществовавший до 823 г. Завоевание мадьярами региона началось в 895 г., завершилось в 900 г. С востока в него пришли «seven Hungarian («Magyar») and three «Kabar» tribes accompanied by fragments of other ethnic groups of eastern origin» [Ery, 1983, с. 105]. Правила мадьярами династия Арпадов (889-1301 гг.). В XIII в. на территорию Венгрии (с 1000 г. королевство) было вторжение монголов. В этот период на Среднедунайскую равнину пришло и здесь осело несколько сообществ.
Среди современных венгров выделяется восемь этнических групп [Guglielmino, 2000]. Группа Orseg локализована около юго-западной границы Венгрии в условиях генетической изоляции со времени прихода мадьяр в конце IX в. Группа Paloc – на севере Венгрии. Говорит на специфическом венгерском диалекте. Регион проживания группы был оккупирован аварами в VI в. Группа Matyo локализована восточнее Paloc. В свидетельствах 1275 г. регион упоминался как «необитаемый», но около 1500 г. он заселен жителями сопредельных областей. Сегодня Matyo является изолированной группой. Это, возможно, связано с тем, что ее представители католики, а жители соседних областей – протестанты. Группа Szekely (Seklers) локализована в Трансильвании (Румыния) в Буковине. Секлерыпоселилисьтам в XI-XIIвв. Они отразили атаки печенегов (Pecheneng) и куманов (Kun) поскольку их мужчины были воинами (they were mainly soldiers). Группа Csango (Chango) локализована в Молдавии (Румыния) в регионе, который назывался Etelkoz. Это одна из групп мадьяр, осевших там во время завоевания ими Среднедунайской равнины. По другой гипотезе они поселились в Молдавии после монгольских завоеваний XIII в. Группы Kiskun и Nagykun (их еще называют Small and Great Cumanians (Kuns)) считают своих предков тюрками, которые пришли на территорию Венгрии во время монгольского вторжения 1241-1242 гг. Эти две группы разделены Тисой. Группа Jasz (Yazigian) пришла в Венгрию вместе с куманами (Kuns) тоже во время вторжения. Она имеет иранское происхождение и до XVI в. говорила на иранском языке.
В публикации [Guglielmino, 1996] приводятся некоторые уточняющие сведения по этническим группам венгров. Предки Kiskun и Nagykun до прихода в Венгрию жили в западной части Центральной Азии и говорили на тюркском языке. Кальвинизм (христианство протестантского толка) они приняли в XVI в. Jasz пришли в Венгрию после монгольского вторжения («They arrived in Hungary after the Tatar invasion (1241-1242)»). Они приняли католическую форму христианства.
3. Аутосомные маркеры
Выборка авторов публикации [Guglielmino, 2000] включала аутосомные маркеры восьми этнических групп венгров, венгров Будапешта, а также юго-запада и севера Венгрии, итальянцев, немцев, поляков и славян (сербов, болгар, чехов и русских). В неё включены популяции, предки которых могли быть родственны мадьярам. Выборка обработана методом главных компонент. На соответствующей диаграмме нами выделено три кластера [Тюрин, 2000, Венгры, рис 2]. В кластер «Европейцы» попали точки «Немцы», «Итальянцы», «Славяне», «Поляки», а также «Венгры Будапешта» и «Венгры юго-запада Венгрии». Точка «Венгры севера Венгрии» в этот кластер не попадает, но расположена логично по отношению к его структуре.
В кластер «Этнические группы венгров» I попали точки «Nagykun» (тюркский язык), «Paloc» и «Orseg» (венгерский язык), а также «Jasz», (иранский язык). Первые три точки кластера характеризуют группы венгров, живущих на северо-востоке Венгрии, последняя – в ее западной части. В кластер «Этнические группы венгров» II попали точки «Szekely», «Csango» и «Matyo» (венгерский язык), а также и «Kiskun» (тюркский язык). Две точки, характеризующие этнические группы венгров, говоривших на тюркском языке – «Kiskun» и «Nagykun», попали в разные кластеры. Точки «Szekely» и «Csango» практически наложились друг на друга.
Таким образом, венгры в целом не отличаются от европейцев. По всем их этническим группам вывод противоположный. Вместе с тем, их близость к каким-либо популяциям Евразии не просматривается. Этнические группы Paloc, Orseg, Kiskun и Jasz, предки которых говорили на венгерском, тюркском и иранском языках генетически близки. Это означает, что их предки входили в одно и то же сообщество, части которого пришли на Среднедунайскую равнину и фигурировали в свидетельствах под разными этнонимами. Это же относится и к группам Szekely, Csango, Kiskun и Matyo.
4. Гаплогруппы Y-хромосомы венгров, мадьяр, аваров и гуннов
4.1. Венгры
Нами найдено шесть массивов гаплогрупп Y-хромосомы, характеризующих венгров. Они сформированы генетиками в научных целях: [Tambets, 2004] (N=113), [Semino, 2000] (N=45), [Csanyi, 2008] (N=197), [Battaglia, 2008] (N=53), [Volgyi, 2008] (N=119), [Pamjav, 2017] (N=147). Всего N=674. Выявлен 1 носитель гаплогруппы С. В базе гаплотипов и гаплогрупп [FamilyTreeDNA, 2017] (N=500) тоже 1 носитель этой гаплогруппы. У секлеров [Клёсов, 2009] (N=143) и [Brandstatter, 2007] (N=97) носители монгольских гаплогрупп не выявлены. Формальный вывод здесь однозначный. На территорию Венгрии какие-либо сообщества, имеющие высокий процент носителей «монгольских» гаплогрупп, не приходили. Другими словами, не приходили сообщества, сформированные в Центральной Азии.
Номенклатуры субкладов гаплогруппы N менялись несколько раза. В нашем тексте сохранены их обозначения, применяемые авторами публикаций. Авторы публикации [Post, 2019] изучили субклады этой гаплогруппы. Установили, что линия B540 субклада N3a4-Z1936, является общей для венгров, хантов и манси, а линия B545 – венгров, башкир и поволжских татар. Субклады гаплогруппы N башкир изучены по массиву данных, приведенных в публикации [Лобов, 2009]. Среди их географически обособленных субпопуляций частота гаплогруппы распределена крайне неравномерно – от 1,4 % в Баймакском районе (N=74) до 64,7 % на востоке Оренбургской области (N=34) [Post, 2019]. У последних и самая высокая частота носителей субклада N3a4-Z1936 – 58,8 %. Он представлен только линией B540. У хантов (N=53) и манси (N=96) ее частоты 7,5 и 19,8 % соответственно. У башкир Стерлитамакского района (N=54) гаплогруппа N представлена только линий B545 – 51,9 %. У татар (N=240) представлены обе линии субклада: B540 – 4,2 %, B545 – 1,3 %. У эстонцев (N=279), латвийцев (N=86), литовцев (N=111), белорусов (N=267), поляков (N=93) русских (N=677), казахов (N=185) и узбеков (N=215) носителей рассматриваемого субклада не имеется. У популяций Южной Сибири распространена линия N3a родительская по отношению к субкладу N3a4-Z1936. Ее частоты у хакасов-сагайцев (N=69) 36,2 % [Дамба, 2018]. Всего частота гаплогруппы N 63,8 %. Причем у них не выявлены «монгольские» гаплогруппы. У тувинского рода ооржак (N=27) частота линии N3a 29,6 %, гаплогруппы N 59,3 %, у рода монгуш (N=64) – 9,4 и 39,1 % соответственно. У монголов (N=852) и алтайцев (N=76) линии N3a не имеется.
Ниже приведены фактические данные по венграм. В массиве [Tambets, 2004] 1 носитель гаплогруппы N (0,9 %) – субклад N3a4-Z1936. В массиве [Pamjav, 2017] – 9 (6,1 %), в том числе 2 (1,4 %) имеют субклад N3a4-Z1936. В [FamilyTreeDNA, 2017] их 14 (2,8 %) и 7 (1,4 %) [Post, 2019] соответственно. Всего (N=760) – 24 (3,2 %) и 9 (1,2 %). Исходя из этого, 1,2 % венгров имеют общих предков с хантами, мансами, а также популяциями Южного Урала и Волго-Уральского региона. Секлеры охарактеризованы двумя массивами данных. В массиве [Клёсов, 2009] 3 носителя (2,1 %) гаплогруппы N, в массиве [Brandstatter, 2007] (N=97) – 1 (1 %). Но это без идентификации субкладов. В массиве из публикации [Post, 2019] (N=95) их 6 (6,3 %), в том числе 4 (4,2 %) носителя субклада N3a4-Z1936. В соответствии с последним массивом у секлеров частоты гаплогруппы N и ее рассматриваемого субклада явно выше, чем у венгров в среднем. Причем, субклад N3a4-Z1936 представлен только линией B540, характерной для хантов и манси.
Выводы авторов публикации [Post, 2019] крайне осторожные до невнятности: «However, a limited portion of specific Y-chromosomal lineages from haplogroup N, sometimes associated with the spread of Uralic languages, link modern Hungarians with populations living close to the Ural Mountain range on the border of Europe and Asia». Высокие частоты определенных линий гаплогруппы N действительно связаны с популяциями, говорящими на языках уральской семьи. Действительно у венгров имеются ее линии, которые связывают их с популяциями, локализованными вблизи Уральских гор. Наши же выводы вполне определенные. На территорию Венгрии в прошлом пришло две группы переселенцев. Предки одной из них были локализованы на юге Западной Сибири, в Южном Зауралье на территории Северного Казахстана. Там они получили от предков башкир востока Оренбургской области линию B540 субклада N3a4-Z1936. Этническим компонентом секлеров является только эта группа. Локализация предков венгров второй группы – Волго-Уральский регион и Южный Урал. Там они получили обе линии рассматриваемого субклада. Но не исключается, что родина переселенцев была в Южной Сибири (носители линии N3a). Оттуда они пришли в обозначенные регионы и на Среднедунайскую равнину.
4.2. Мадьяры, авары и гунны
В публикации [Neparaczki, 2019] приведены результаты тестирования останков из погребений гуннов (V в.), аваров (конец VI – VII вв.) и мадьяр (конец IX – X вв.). По образцам, характеризующим мадьяр (N=30), получены следующие гаплогруппы и субклады. Гаплогруппа R1a (6 образцов) представлена двумя субкладами – R1a1a1b1a2b-CTS1211 (4 образца) и R1a1a1b2-Z93 (2 образца). Популяции с высокими частотами субклада R1a1a1b1a2b-CTS1211: словенцы – 29,7 (6,7), словаки – 26,5 (15,0), литовцы – 24,6 (12,3), русские – 23,9 (19,2), поляки – 23,3 (32,7), хорваты – 20,6 (3,4), венгры – 19,8 (8,8), эстонцы – 19,7 (9,8), белорусы – 19,6 (30,1), боснийцы и герцеговинцы – 16,2 (1,2), украинцы – 16,1 (26,2) % [Haplogroup R1a]. В скобках указаны суммарные частоты субкладов R1a-M458 и R1a-Z92. Субклад R1a1a1b1a2b-CTS1211 не мог избирательно попасть к мадьярам от восточных и западных славян. Вместе с ним к ним попали бы и другие характерные для них субклады гаплогруппы R1a. Остаются южные славяне с относительно низкими частотами двух «не мадьярских» субкладов. Субклад R1a1a1b2-Z93 был распространен у популяций и групп степной и лесостепной зон Евразии [Narasimhan, 2019]. Сегодня его относительно высокие частоты у поволжских татар, кубанских ногайцев, караногайцев, киргизов, алтайцев. Этот субклад обязан был быть у мадьяр.
Гаплогруппа R1b (5 образцов) представлена субкладами R1b1a1b1a1a1-U106 (4 образца) и R1b1a1b1a1a2b-U152 (1 образец). Первый распространен в Германии, Скандинавии и Британии. С небольшими частотами в Восточной Европе. Наибольшие частоты второго субклада в районе Альп.
Гаплогруппа I2 (6 образцов) представлена субкладом I2a1a2b-L621 (5 образцов). Наибольшие его частоты на Балканах. С небольшими частотами распространен в Восточной Европе. Скорее всего, оставшийся образец имеет этот же субклад.
Гаплогруппа N (5 образцов) представлена двумя субкладами. Субклад N1a1a1a1a2-Z1936 (3 образца) имеет высокие частоты у саамов, финнов и карелов, распространен у популяций Волго-Уральского региона и башкир. Имеется у караногайцев, сибирских татар, хантов и манси. Субклад N1a1a1a1a4-M2118 (2 образца) доминирует у якутов. Его небольшие частоты у хантов, манси и казахов.
Гаплогруппа E (2 образца) представлена субкладом E1b1b1a1b1a-V13 (1 образец). Скорее всего, второй образец тоже имеет этот субклад. Высокие частоты субклада на Балканах. По 5 образцам определены гаплогруппы I1, J1, J2a1a, G2a2b и Q1a. По неопубликованным данным по одному образцу, характеризующему мадьяр, определена гаплогруппа C. Мы его учли в общем количестве тестированных останков. Гаплогруппа I1 со значимыми частотами распространена у русских и популяций Средней Волги. Гаплагруппа G2a2b определена у останков, отнесенной к баденской культуре (3600-2800 гг. до н. э.) на территории Венгрии [Narasimhan, 2019]. То есть, она в регионе является реликтовой. Можно принять, что гаплогруппы J1 и J2a1a являются северокавказскими, а локализация гаплогрупп Q1a неопределенная. Но, скорее всего, ее «родина» в Восточной Евразии.
Гаплогруппы и их субклады, локализованные в Западной Европе и на Балканах – 19 (63,3 %), условно на Средней Волге – 6 (20 %), на Северном Кавказе – 2 (6,7 %), в Восточной или южной Сибири – 2 (6,7 %), Центральной Азии – 1 (3,3 %). Итого, среди мадьяр имеется 36,7 % потомков тех, кто пришел на Среднедунайскую равнину с востока, в том числе 10,0 % – из Восточной или Южной Сибири и Центральной Азии.
У аваров (N=14) доминирует гаплогруппа N (7 образцов). Еес убклад N1a1a1a1a3-B197 (5 образцов) имеет высокие частоты у бурятов, чукчей, эскимосов, коряков. С невысокими частотами распространён у тувинцев и монголов. Другие два образца могут иметь этот же субклад. Гаплогруппа R1a (2 образца) представлена линией R1a1a1b2a-Z94 субклада R1a1a1b2-Z93. По 2 образцам определена гаплогруппа С. По трем – гаплогруппы I1, G2a и E1b1b1a1b1a. Формально предки аваров локализуются в Восточной Сибири и Центральной Азии. Из этих регионов все носители гаплогрупп N и С (суммарно 64,3 %). Но, скорее всего, в Туве. У тувинцев (N=936) 35,9 % носителей гаплогруппы N, в том числе 8,9 % ее родительской линии по отношению к субкладу N1a1a1a1a3-B197 и 14,3 % носителей гаплогруппы С [Харьков, 2013]. Имеются 13,0 % носителей линии R1a1a (M17) родительской по отношению к R1a1a1b2a-Z94.
По образцам, характеризующим гуннов (N=3), получены гаплогруппы R1a1a1b2a2, R1b1a1b1a1a1 и Q1a2.
Определить соотношение субклада N3a4-Z1936 современных венгров с субкладами мадьяр и аваров мы не смогли. По аутосомным маркерам с высокой вероятностью цвет глаз у аваров и гуннов был коричневый, а волос – темно-коричневый и черный. Но у одного авара – светлый (блондин). Аналогичная информация по мадьярам приведена ниже.
Методом главных компонент (MDS) обработаны генетические расстояния между популяциями, определенные по частотам гаплогрупп Y-хромосомы. Общая интерпретация диаграммы популяций Евразии (N=78) [Neparaczki, 2019, fig. 2] однозначная. По ее горизонтальной оси слева направо возрастает уровень центрально-азиатской монголоидности, а по вертикальной снизу-вверх – уралоидности. Точка «Мадьяры» находится между точками «Русские» и «Финны», но экстремально близко к первой. Рядом точка «Туркмены». На некотором удалении от точки «Мадьяры» точки «Селькупы», «Кеты» и «Восточная Европа». Точка «Авары» находится далеко от других точек. Справа от нее «Якуты», «Долгане» и «Коряки». Снизу – «Казахи» и «Киргизы». Слева – «Телеуты», «Коми», «Ханты», «Ненцы» и «Нганасане». Дать общую интерпретацию диаграммы выборки популяций Европы и Сибири (N=58) [Neparaczki, 2019, fig. 3] мы не можем. Она включает только мадьяр. Их точка практически совпала с точкой «Башкиры». Точки других популяций находятся от них далеко.
Общий вывод авторов публикации [Neparaczki, 2019, р. 1]: «Composition of conquering Hungarian paternal lineages is very similar to that of Baskhirs, supporting historical sources that report identity of the two groups». Добавим, что генофонд мадьяр по гаплогруппам Y-хромосомы и их субкладам похож также и на генофонд русских. Но это мы не примем во внимание, поскольку субклад R1a1a1b1a2b-CTS1211 не мог попасть к ним от русских.
5. Гаплогруппы мтДНК венгров, мадьяр и аваров
5.1. Венгры
По двум выборкам у венгров выявлено 3-5 % восточноевразийских гаплогруппмт ДНК [Maroti, 2018]. Авторы последней публикации выполнили анализ гаплогрупп венгров своей выборки (N=272). Среди них восточноевразийских менее 3 %. Главный вывод: «the majority of modern Hungarian maternal lineages have Late Neolith/Bronze Age European origins (partially shared also with modern Danish, Belgian/Dutch and Basque populations), and a smaller fraction originates from surrounding (Serbian, Croatian, Slovakian, Romanian) populations». У поляков и чехов (N=708) 1,8 % восточноевразийских гаплогрупп, у южных славян (N=1705) – 1,2 %, у русских (N=526) – 1,3 % [Malyarchuk, 2006]. В публикации [Brandstatter, 2007] приведены данные по двум этническим группам венгров – секлерам (N=178) и Csango (N=182). По набору и частотам гаплогрупп они практически не различаются. Восточноевразийских гаплогрупп у секлеров 8 % у Csango 6 %. Это явно выше, чем у венгров в целом, однако, соответствует их доле у румын исторических областей Румынии: Валахия (N=226) – 5,8 %, Добруджа (N=466) – 4,3 %, Молдавия (N=235) – 7,2 %, Трансильвания (N=208) – 2,9 % [Cocos, 2017]. Всего у румын (N=715) – 5,3 %.
5.2. Мадьяры и авары
В публикации [Csosz, 2016] приведены результаты обработки генетических расстояний, определенных по частотам гаплогрупп мтДНК выборки древних сообществ. Онавключаетмадьяр (N=76) иаваров (N=26). Точки «Мадьяры», «Бараба» и «Минуса» находятся экстремально близко друг к другу. Вторая характеризует останки в погребениях позднего бронзового века в Барабинской степной зоне, третья – бронзового века в Минусинской впадине. Точка «Авары» находится недалеко от этих точек. Недалеко от нее точки «Викинги на территории Дании» и «Польша» (Средние века). У аваров выявлено 4 (15,4 %) восточноевразийские гаплогруппы. При кластеризации авары и поляки образовали один изолированный кластер. То есть, жены аваров «тувинцев» были в основном западными славянками.
По останкам в восьми погребениях на территории Венгрии, отнесенных к мадьярам, определены гаплогруппы мтДНК (N=102) [Neparaczki, 2018] (включают и те, которые приведены в публикации [Csosz, 2016]). Среди них 31 (30,4 %) отнесены к восточноевразийским, том числе и субклады U2e1b (3 образца) и T2b4h (1 образец), что не соответствует выделению этой ассоциации гаплогрупп российскими специалистами [Лобов, 2009]. С учетом этого, частота восточноевразийских гаплогрупп у мадьяр составляет 26,5 %. Вывод здесь однозначный. На территорию Венгрии в прошлом пришло сообщество (пришли сообщества) с аномально высокими для этого региона частотами восточноевразийских гаплогрупп.
Выполнено сопоставление восточноевразийских гаплогрупп мадьяр с другими массивами данных. Высокий компонент их генофонда (26-41 %) выявлен у поволжских татар. Возможность прихода их предков – булгар, на территорию Венгрии подтверждается археологическими и антропологическими данными, а также историческими свидетельствами. Отмечено, что мадьяры и булгары имели схожие антропологические характеристики («characterized by similar anthropological types» [Neparaczki, 2018]).
Гаплогруппы мадьяр авторы разделили на «субпопуляции» European, European+Eurasian, EastEurasian и EastEurasian+Eurasian, выполнили их сопоставление с гаплогруппами, характеризующими современные популяции и представителей археологических культур. Генетические расстояния между ними обработали методом главных компонент (Рис. 1).Точки European и European+Eurasian практически совпали. Это свидетельствует о том, что «субпопуляция» Eurasian тождественна European. Обе практически тождественны представителям срубной культуры (кластер III). «Субпопуляция» EastEurasian тождественна тувинцам (кластер I). У последних (N=36) 72,3 % восточноевразийских гаплогрупп [Деренко, 1999]. Микс «субпопуляций» EastEurasian+Eurasian не соответствует каким-либо популяциям и группам. Это еще раз свидетельствует о том, что «субпопуляция» Eurasian выделена некорректно. В целом мадьяры экстремально близки европейским скифам, представителям потаповской и полтавкинской культур, а также татарам (кластер II) и максимально далеки от венгров. Рядом с кластером II находятся русские, популяции Кавказа и Северной Европы.
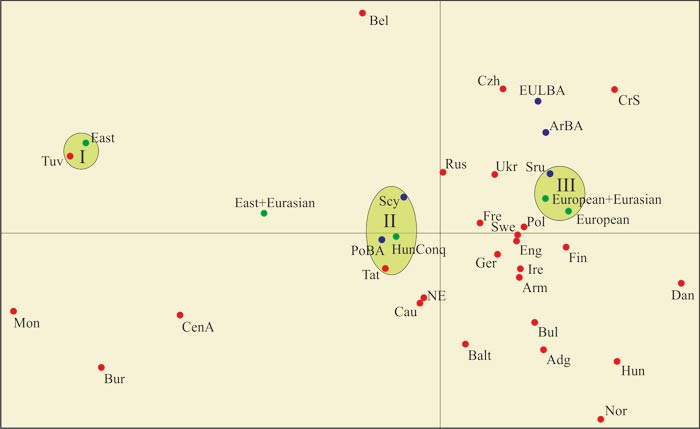
Рис. 1 – Диаграмма главных компонент (multidimensional scaling) гаплогрупп мтДНК современных популяций и представителей археологических культур [Neparaczki, 2018, fig. 4]. Кластеры выделены А.М. Тюриным
Археологические культуры (синий кружок): ArBA– ?; EULBA – поздний бронзовый век Европы; Scy – европейские скифы; PoBA – потаповская и полтавкинская культуры; Sru – срубная культура Понто-Каспийского региона; мадьяры (зеленый кружок): HunConq – весь массив гаплогрупп. «Субпопуляции» мадьяр на диаграмме подписаны.
Современные популяции: Adg – адыги; Arm – армяне; Balt– популяции Прибалтики; Bel – белорусы; Bul – болгары; Bur – буряты; Cau – популяции Кавказа; CenA – популяции Центральной Азии; CrS – хорваты и сербы; Czh – чехи; Dan – датчане; Eng – англичане; Hun – венгры; Ger – немцы; Fin – финны; Fre – французы; Ire – ирландцы; NE – популяции Северной Европы (?); Nor – норвежцы; Mon – монголы; Pol – поляки; Rus – русские; Swe – шведы; Tat – повольжские татары; Tuv– тувинцы; Ukr – украинцы.
Потаповская культура (бронзовый век, первая четверть II тыс. до н. э.) локализована в лесостепной части Волго-Уральского региона. Полтавкинская (бронзовый век, 2700-2100 гг. до н. э.) – в Среднем Поволжье и по долине реки Самара. Европейские скифы (ранний железный век, VIII в. до н. э. – IV в. н. э.) – в Северном Причерноморье. Это четко локализует кластер II. Северное Причерноморье и Волго-Уральский регион, включая территорию Татарстана.
У скифов Северного Причерноморья (N=19) 26,3 % восточноевразийских гаплогрупп мтДНК [Juras, 2017]. В статье [Конюхова, 2010] приведены частоты гаплогрупп казанских татар (N=135), мишарей (N=140) и крещеных татар (N=84). У казанских татар 23,0 % восточноевразийских гаплогрупп, у мишарей – 20,0 %, у крещеных татар они не выявлены. Сам по себе это интереснейший факт. Но мы отметим только то, что в целом (с учетом небольшой численности последних) доля восточноевразийских гаплогрупп у татар примерно соответствует таковой у мадьяр. Русские и поволжские татары по набору и частотам гаплогрупп мтДНК характеризуются минимальным генетическим расстоянием. [Naumova, 2009]. Это свидетельствует о том, что западноевразийская составляющая генофонда мадьяр близка к генофонду русских.
По гаплогруппам Y-хромосомы и мтДНК мужчин мадьяр (N=29) двумя способами («Multinomial logistic regression average classification» и «Naive Bayes (H-W) predicted admixture») оценена вероятность происхождения их предков в Европе, Восточной Азии и Африке [Neparaczki, 2018]. По первому способу с вероятностью более 90 % предки 10 локализованы в Европе, 6 – в Восточной Азии, по второму – 20 и 6 соответственно. При учете одного мадьяра, носителя гаплогруппы С, у мадьяр минимум 22,6 % предков из Восточной Азии. По аутосомным маркерам с высокой вероятностью цвет глаз у мадьяр, имеющих предков из Европы, был коричневым и голубым, из Восточной Азии – коричневым. Цвет волос первых – светло- и темно-коричневым, один был рыжим, один блондином. У вторых – темно-коричневым и черным.
6. Апарды
В публикации [Olasz, 2019] приведены результаты тестирования останков венгерского короля Белы III (династия Арпадов, 1172-1196 гг.), его супруги Анны Антиохийской и еще восьми человек, найденных в королевской базилике. У Белы III гаплогруппа Y-хромосомы R1a, мтДНК – H1b, у Анны – H1j8/H1bz. У другой женщины – T. Гаплогруппы других мужчин R1a (2), R1b (2), J1 и E1b1. У людей, чьи останки погребены в базилике, восточноевразийских гаплогрупп не имелось. Для мужчин определены 17-маркерныегаплотипы, а по ним – гаплгруппы. Один носитель гаплогруппы R1a близкий родственник короля. Мы выполнили сравнение гаплотипов носителей гаплогруппы R1a с базовыми гаплотипами ее субкладов. Наиболее вероятный субклад у Белы III R1a-Z93. Заключение А.А. Клёсова (по нашей просьбе): «Это похоже на балто-карпатскую ветвь, R1a-Z280-CTS3402». Отмечена низкая достоверность таких определений по 17-маркернымгаплотипам. Балто-карпатская ветвь субклада R1a-Z280 сегодня распространена, в основном, у западных и восточных славян [Клёсов, 2015].
7. Полногеномные маркеры
На рисунке 2 приведена диаграмма главных компонент полногеномных маркеров [Järve, 2019]. На ней современные популяции (медианы точек полногеномных маркеров) и индивидуальные точки, характеризующие останки представителей археологических культур. Современные популяции локализованы на диаграмме на удивление логично. По положению характеризующихих точек четко обозначились два потока генов. Первый – монголоидность из Центральной Азии до территорий Турции и Сирии, второй – уралоидность из Восточной Сибири до территории Эстонии. Более подробно наша интерпретация диаграммы приведена в публикации [Тюрин А.М., 2019, Скифы]. Здесь для нас важно то, что венгры не имеют ни малейших компонентов монголоидности и уралоидности. Не отличимы от словенцев, близки немцам и боснийцам. Скифы Венгрии (N=5) европеоиды без монголоидных и уралоидных компонентов. Один скиф – грек, один – македонец, румын или болгарин, один – босниец, два – словаки или немцы. Вполне возможно, три последних скифа попадают в «облако» венгров. Скифы Венгрии не являются популяцией.
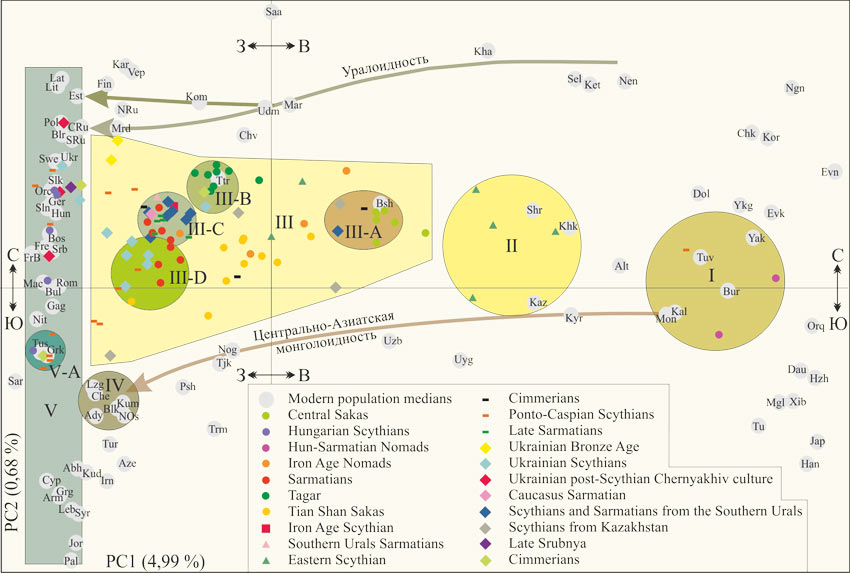
Рис. 2 – Диаграмма главных компонент полногеномных маркеров современных популяций Евразии (показаны медианы) и древних геномов [Järve, 2019]. Интерпретация (кластеры и стрелки потоков генов) А.М. Тюрина
Европа: Bsh, башкиры; Blr, белорусы; Bos, боснийцы; Bul, болгары; Chv, чуваши; Cyp, киприоты; Est, эстонцы; Fin, финны; Fre, французы; FrB, баски Франции; Gag, гагаузы; Ger, немцы; Grk, греки; Hun, венгры; Kar, карелы; Kom, коми; Lat, латвийы; Lit, литовцы; Mac, македонцы; Mar, марийцы; Mrd, мордва; Nit, северные итальянцы; Orc, оркнейцы; Pol, поляки; Rom, румыны; NRu, северные русские; CRu, центральные русские; SRu, южные русские; Saa, саамы; Sar, сардинцы; Srb, сербы; Slk, словаки; Sln, словенцы; Swe, шведы; Ttr, татары; Tus, тосканцы; Udm, удмурты; Ukr, украинцы; Vep, вепсы.
Сибирь: Alt, алтайцы; Bur, буряты; Chk, чукчи; Dol, долгане; Evk, эвенки; Evn, эвены; Ket, кеты; Khk, хакасы; Kha, ханты; Kor, коряки; Mon, монголы; Nen, ненцы; Ngn, нганасаны; Sel, селькупы; Shr, шорцы; Tuv, тувинцы; Yak, якуты; Ykg, юкагиры.
Восточная Азия: Dau, даурцы; Han, китайцы; Hzh, нанайцы (Hezhen); Jap, японцы; Mgl, монголы; Orq, орогены; Tu, Tu; Uyg, уйгуры; Xib, Xibo.
>Центральная Азия: Irn, иранцы; Kaz, казахи; Krd, курды; Kyr, киргизы; Psh, пуштуны; Tjk, таджики; Trm, туркмены; Uzb, узбеки.
Ближний восток: Jor, иорданцы; Leb, ливанцы; Pal, палестинцы; Syr, сирийцы; Tur, турки.
Кавказ: Abh – абхазы; Ady – адыгейцы; Arm – армяне; Aze – азербайджанцы; Blk – балкарцы; Che – чеченцы; Grg – грузины; Kal – калмыки; Kum – кумыки; Lzg – лезгины; Nog – ногайцы; Nos – северные осетины.
8. Антропология
В публикации [Neparaczki, 2018] приведены примеры европеоидного и европеоидно-монголоидных черепов мадьяр (Рис. 3). По нашему мнению, российские антропологи идентифицировали бы два последних как европеоидные. Автор рецензии [Козинцев, 1972] указал на то, что зарубежные исследователи начала 30-х годов прошлого века и позднее при анализе древнего краниологического материала с территории Венгрии «исходили из весьма устаревших представлений об антропологических особенностях монголоидов» (с. 166). Но и с момента ее публикации представления специалистов в этом вопросе эволюционировали. Исходя из этого, констатируем, что в выделении монголоидных краниологических характеристик имеются проблемы. Поэтом не примем во внимание заключения авторов публикации [Neparaczki, 2018] по черепам мадьяр. Ниже рассмотрены только сравнительные характеристики краниологических серий, обработанных формальными методами.
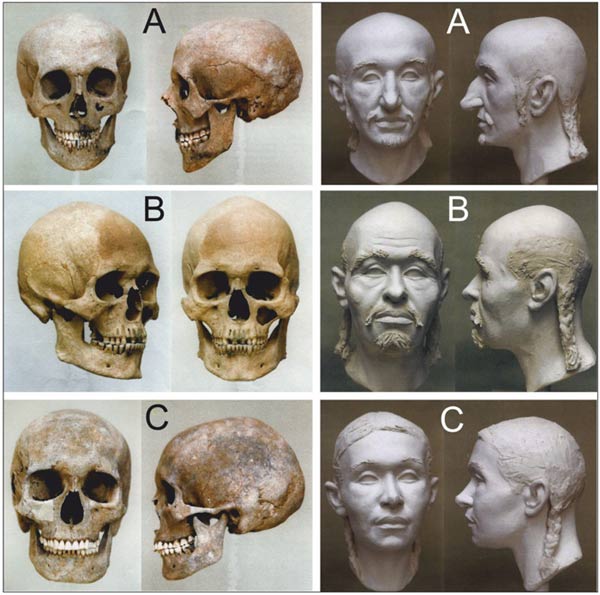
Рис. 3 – Черепа мадьяр и реконструкции облика лица [Neparaczki, 2018, fig. 6]. Мужчина европеоид (A), мужчина (B) и женщина (C) с европеоидно-монголоидными краниологическими характеристиками
Автор публикации [Тот, 1977] обработал методом главных компонент краниологические серии (мужские и женские черепа), характеризующие представителей археологических культур, включая мадьяр. На диаграмме энеолита и бронзового века рядом с точкой «Мадьяры» находятся токи «Андроновская культура» (Восточный Казахстан) и «Сиалк» (Иран). На диаграмме VII-I вв. до н. э. – «Савроматы» (Нижнее Поволжье) и «Тагарская культура I» (Минусинская впадина), на диаграмме I-VIII вв. н. э. – «Средние сарматы» (Калиновка, Нижняя Волга), «Сарматы» (Украина) и «Миздахкхан» (Приаралье).
Выборка другого автора [Ery, 1983] включает 45 краниологических серий (мужские черепа) из Карпатского бассейна (Среднедунайская равнина) и 75 серий (железный век и средневековье) из Южной Сибири, Тян-Шаня, района Арала, Центральной и Восточной Европы (степная и лесостепная зоны). Аварский период (VI-VIII вв.) Карпатского бассейна характеризуют 22 серии. По географическому признаку они разделены на четыре группы. По погребениям первых трех генераций завоевателей (Period of the Hungarian Conquest, 900-980 гг.) сформированы 4 группы (A, D, C, D). В 1000-1301 гг. в Венгрии правила династия Арпадов. Этот период охарактеризован 18 сериями. По географическому признаку они разделены на пять групп. Таким образом, предки венгров охарактеризованы 13 сериями. Другие серии объединены в 47 групп. Они включают четыре группы аварского периода и две арпадского. Рассчитаны генетические расстояния между краниологическими характеристиками групп и выполнена их кластеризация. Нами выделены кластеры, включающие аваров, мадьяр и арпадов (Рис. 4).
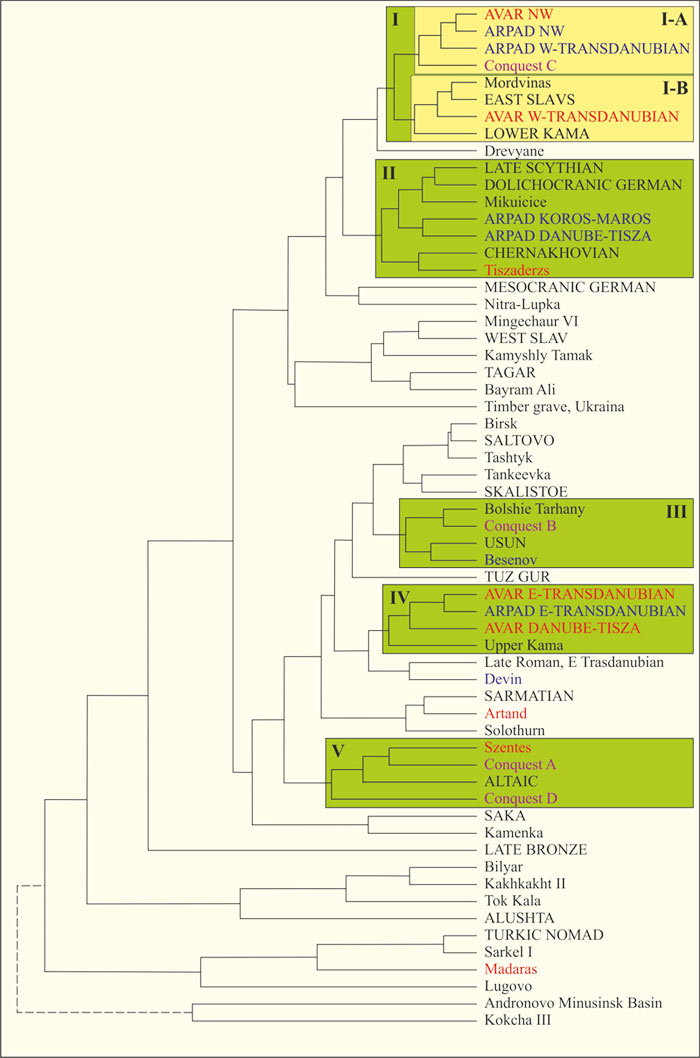
Рис. 4 – Дендрограмма генетических расстояний между краниологическими характеристиками групп представителей археологических культур степной и лесостепной зон Евразии (мужские черепа) [Ery, 1983, fig. 82]. Цветом показаны группы аваров и соответствующего им периода (красный), мадьяр (малиновый) и арпадов и соответствующего им периода (синий). Кластеры выделены А.М. Тюриным
В подкластер I-А попали только венгерские группы. Минимальные генетические расстояния между арпадами и аварами северо-запада Венгрии. Это означает, что по краниологическим характеристикам они практически неразличимы. Близки к ним арпады из региона к западу от Дуная, а также мадьяры группы С. Последние локализованы в основном к западу от Дуная.
Авары из региона к западу от Дуная попали в подкластер I-B. В него попали группы Mordvinas, EAST SLAVS и LOWER KAMA. Первая включает черепа из Муранского грунтового могильника (Самарская лука, правый берег Волги, относится к сельским могильникам Волжской Булгарии) и из могильника Tsna (?) (XIII-XIV вв. н. э.). Вторая – дреговичей, полян, северян и радимичей. Группа LOWER KAMA – могильник Мазунинский и Чегантинскую группу могильников (II в. до н. э. – VI в. н. э.). Черепа из Мазунинского могильника имеют явные признаки монголоидности: «широкое лицо, сравнительно высокие орбиты, слабое выступание носа и более низкое переносье» [Акимова, 1961, с. 123]. Для Чегандинской группы характерны черепа двух типов: «европеоидный и смягченно монголоидный» (с. 122). На диаграмме главных компонент, точка, характеризующая Муранский могильник, находится за пределами «облака», точек могильников Волжской Булгарии [Руденко, 2016, рис. 12]. То есть, погребенные в нем не характеризуют булгар. Исходя из этого, кластер I следует идентифицировать как восточнославянский, включающий и группы Среднего Поволжья. При такой кластеризации последние являются славянами с монголоидными и уралоидными компонентами. Этот кластер включает пять групп предков венгров.
Две группы арпадов и могильник Tiszaderzs аварского периода попали в кластер II. Он также включает поздних скифов, долихокранных германцев, западных славян (Mikuicice) и черняховцев. Черняховцев мы однозначно идентифицировали как «славяне» [Тюрин, Готы, 2019; Скифы, 2019]. Причем в один кластер с ними попали и краниологические серии поздних скифов [Тюрин, Готы, 2019, рис 6]. Кластер II можно идентифицировать как западнославянский. Отметим, что по краниологическим характеристикам поздних скифов Северного Причерноморья (N=323) сделано предположение, что они имеют местные и центральноазиатские компоненты [Movsesian, 2017]. Но доля и происхождение последних не определены. Естественно, авторы последней публикации рассматривали более представительную выборку серий скифских черепов, чем та, которой оперировал автор публикации [Ery, 1983]. Возможно, в ней имелись и серии с признаками монголоидности.
Кластеры I и II объединены в мегакластер. В него попали и древляне Украины. Мегакластер идентифицируется как славянский (восточные и западные славяне).
Мадьяры группы D (северо-восток Венгрии) попали в кластер III. Он включает могильник арпадского периода Besenov, Больше-Тарханский могильник (правый берег Волги, Республика Татарстан, относится к сельским могильникам Волжской Булгарии), и усуней (Казахстан и Киргизия, III в. до н. э. –I в. н. э.). На диаграмме главных компонент, точка, характеризующая Больше-Тарханский могильник, попала в центр «облака» точек группы могильников Волжской Булгарии [Руденко, 2016, рис. 12]. То есть, мадьяры группы D по краниологическим характеристикам близки населению Волжской Булгарии. Это же относится к группе из могильника Besenov. Специалисты отмечают: «На всех болгарских сериях с территории СССР прослеживается общий для них так называемый зливкинский тип – европеоидный с ослабленными монголоидными чертами» [Герасимова, 1987]. Авторы публикации [Исмагулов, 2018] рассмотрели краниологические характеристики усуней Семиречья. Они не отличаются от характеристик других синхронных с ними групп с территории Казахстана. Установлено, что у этих сообществ «доля монголоидных элементов находилась на уровне 25 %, а остальную часть – 75 % составили европеоидные черты». Это соответствует доле восточноевразийской составляющей у мадьяр, определенной по данным популяционной генетики.
Между аварами и арпадами региона к востоку от Дуная минимальное генетическое расстояние. В кластер IV с ними попала и другая группа аваров, а также группа Upper Kama. Последняя включает черепа из Митинского и Деменковского курганных могильников (III-V вв. н. э.). Черепа «из Митинского могильника характеризуются в общем европеоидными чертами в сочетании с довольно низким углом выступания носа [Акимова, 1961, с. 123]. Особенности черепов из Деменковского могильника: «По значительной ширине лица и уплощенности лицевого скелета они как будто имеют тенденцию к большей монголоидности, но, с другой стороны, выступание носа у них сильнее» (с. 124). По нашему мнению, этой характеристике примерно соответствует череп B на рисунке 3. Отметим, что черепа из отмеченных двух могильников в регионе не имеют аналогов.
Группы мадьяр A и D (обе локализованы к востоку от Дуная), а также серия из могильника аварского периода Szentes попали в кластер V. В нем и группа ALTAIC (скифская культура V-IV вв. до н. э.). Краниологический комплекс скифов Алтая (Пазырыкская культура) имеет сходство с современной южносибирской расой, но является более европеоидным [Чикишева, 2000]. Можно предположить, что эти три группы с территории Венгрии имеют наиболее выраженные признаки южносибирской расы.
Генетическое расстояние между группой из погребения аварского периода Artand и сарматами (Западный Казахстан, Нижняя Волга, Южный Урал) не очень большое. Основная часть сарматских серий Волго-Уральского региона и Южного Урала принадлежит европеоидам без признаков монголоидности. Имеются серии с признаками монголоидности [Тюрин, 2018, Монголоидность, cарматы]. «Сарматы Венгрии были крайне смешаны в антропологическом отношении. В материалах присутствуют как чисто европеоидные черепа, так и черепа с легкой монголоидной примесью» [Кириченко, 2013, с. 103]. Другая группа из погребений аварского периода Madaras близка к группам из могильника Саркел I (маленькие курганы, Нижний Дон, IX-X вв. н. э.) и TURKIC NOMAD (Алтай и Минусинская впадина VII-XI вв. н. э.). Краниологическая серия, сформированная по малым курганам Саркела, включает европеоидные и монголоидные черепа [Герасимов, 1949]. Группа Devin арпадского периода неотличима от автохтонного населения Среднедунайской равнины (Late Roman).
Таким образом, авары и арпады семи групп по краниологическим характеристикам неразличимы. Они формируют два обособленных сообщества – кластеры I и IV. В первое входят и мадьяры группы C. Одно сообщество арпадов обособилось (кластер II). В него входит и группа аварского периода. Обособились два сообщества мадьяр (кластеры III и V). В одно из них входит группа аварского периода, в другое – арпадского. Можно утверждать, что авары, мадьяры и арпады – это одно и то же сообщество, имеющее региональные особенности краниологических характеристик. Выделяется четыре субпопуляции. Одна (мегакластер I+II) соответствует восточным и западным славянам. Другая (кластер V) имеет характеристики близкие к южносибирской расе. Две субпопуляции (кластеры III и IV) –европеоиды с элементами монголоидности и уралоидности.
9. Литовские татары
Литовские татары, как субэтнос, сформировались из военного сословия политических образований XV в. – Большой Ногайской орды, Малой Ногайской орды и Крымского ханства, являющихся осколками Улуса Джучи XIII-XIV вв. Обособились в Великом княжестве Литовском в начале XVI в. Субэтнос охарактеризован четырьмя массивами данных популяционной генетики: гаплогруппы Y-хромосомы дворянских родов (N=35), гаплогруппы Y-хромосомы (N=74) и мтДНК (N=79), а также полногеномные маркеры (N=6) научных выборок. Все массивы данных сформированы на территории Белоруссии. Массив дворянских родов включает и представителей из других стран. Ранее мы рассмотрели гаплогруппы Y-хромосомы [Тюрин, 2017, Литовские татары]. По отсутствию «монгольских» гаплогрупп у субэтноса сделан вывод: наличие монголов по мужской линии в предках литовских татар исключается. Такое могло быть только в одном случае – монголы не приходили в Восточную Европу и сопредельные регионы Азии ранее XVI в. По результатам авторской интерпретации данных популяционной генетики [Тюрин, 2010, Генетические следы; 2017, Литовские татары; 2017, Казахские торе; 2017, Ногаи, генетика] и антропологии [Тюрин, 2018, Ногаи, антропология] при учете достоверных археологических фактов [Матюшко, 2011] и исторических свидетельств [Трепавлов, 2016] сделан категорический вывод: калмыки были первой популяцией, которая принесла в Восточную Европу и сопредельные районы Азии центрально-азиатскую монголоидность и ее индикаторы – гаплогруппы Y-хромосомы C, O и D. Выше под монголами и калмыками мы имели в виду именно популяции (относительно замкнутые сообщества). Поэтому наличие у того сообщества, которое пришло на Среднедунайскую равнину, 3 (6,8 %) носителей «монгольской» гаплогруппы С ни в коей мере не опровергает наши выводы. Это сообщество не являлось популяцией.
Автор публикации [Рожанский, 2016] выполнил привязку к регионам гаплогрупп Y-хромосомы литовскихих татар. Гаплогруппы N-Tat, R1a-M458, R1a-M558, R1b-M412 и R1b-M478 – Восточная Европа и Волго-Уральский регион, R1a-Z2125 и Q-M242 – Центральная Азия, G2a-U1, J1-P58, J2aM410 и J2b-M12 – Кавказ и Ближний Восток. Гаплогруппы татар Q1a-M346, R1b-M478 и R1a-Z2125 сходны с гаплогруппами популяций Центральной Азии (киргизы, казахи, узбеки), а гаплогруппы G2a-U1 и J1-P58 – с гаплогруппами кавказских популяций. Это подтверждает исторические свидетельства по формированию субэтноса. Но главное, явно восточноевразийских гаплогрупп Y-хромосомы и их субкладов у литовских татар не имеется. При этом у них выявлено 32,3 % восточно-евразийских гаплогрупп мтДНК [Панкратов, 2014].
Выборка полногеномных маркеров авторов публикации [Pankratov, 2016] включает 87 популяций (Рис. 5, А). В поле главных компонент литовские татары сформировали «плотный» кластер, расположенный между популяциями Центральной и Восточной Европы, Центральной Азии и Сибири. По генетическим расстояниям их ближайшие родственники – поволжские татары, башкиры, кубанские ногайцы, таджики и узбеки.
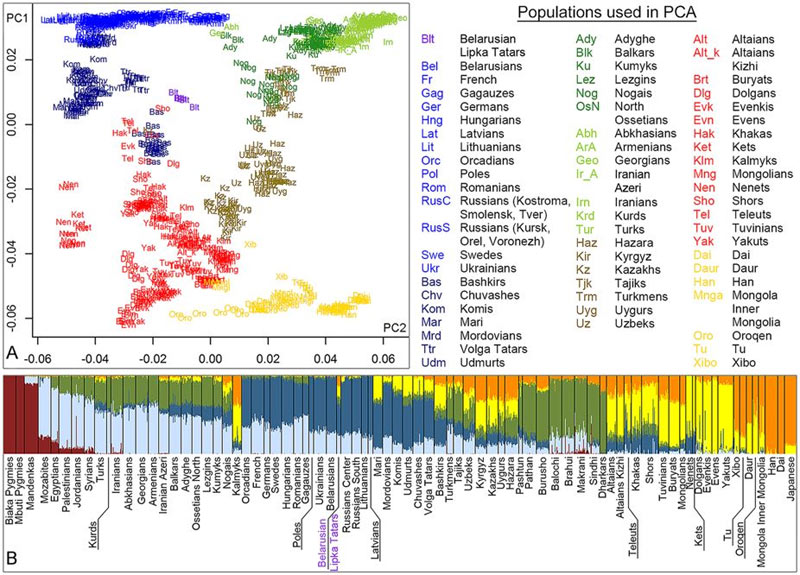
Рис. 5 – Структура полногеномных маркеров популяций Евразии выборки авторов публикации [Pankratov, 2016, fig. 2]. А – диаграммаглавныхкомпонент генетических расстояний между популяциями (PC1 = 3,8, PC2 = 0,6 %). В – результаты обработки полногеномных маркеров популяций методом ADMIXTURE
Источник: https://www.nature.com/articles/srep30197/figures/2
В статье [Yunusbayev, 2015] приведена удачная выборка полногеномных маркеров популяций Евразии, обработанная методом ADMIXTURE. Четко обозначилась ее структура. Выделено восемь кластеров. Они уверенно идентифицируются: Европа (К1), Средний Восток (К2), Кавказ (К3), Южная Азия (К4), Сибирь (К5, К6, К7), Восточная Азия (К8). Маркеры кластеров К6 и К8 можно условно назвать «монгольскими». Они резко доминируют у калмыков – 75 %. У них же заметная доля маркеров кластеров К1 и К3. Это результат метисации калмыков при их контактах, главным образом, с ногаями. Маркеры кластеровК6 и К8 доминируют у киргизов, казахов и каракалпаков. Эти популяции сформированы, в том числе, и на основе калмыков. Заметная доля маркеров кластеров К6 и К8 у популяций Средней Азии – узбеков, таджиков, туркмен, а также кубанских ногайцев. Эти данные явились основой интерпретации полногеномных маркеров литовских татар. У калмыков 82 % «монгольских» маркеров, у кубанских ногайцев – 24 %, у литовских татар – 28 % (Рис. 5, В). На диаграмме «монгольские» маркеры показаны желтым и коричневым цветами.
Возник парадокс. У литовских татар значимо представлены «монгольские» полногеномные маркеры и восточно-евразийские гаплогруппы мтДНК, но не имеется явно восточноевразийских гаплогрупп Y-хромосомы. Второй парадокс отметил автор публикации [Рожанский, 2016]. Это соотношение субкладов R1a-М458 (19 образцов) и R1a-Z280 (5 образцов). Первый имеет высокие частоты у западных славян, второй у восточных. У белорусов частоты этих субкладов 15 и 35 % соответственно. То есть, эта ассоциация двух субкладов не могла попасть к литовским татарам от белорусов. По нашему мнению, здесь все просто. Религии в привычном нам виде начали формироваться только в XVI в. А в XV в. польская шляхта вполне могла наниматься на службу в татарские отряды, постепенно переходя в мусульманство. По частотам у поляков доминирует субклад R1a-М458.
Сегодня литовские татары проживают в Белоруссии (7,3 тыс.), Литве (3,2 тыс.) и Польше (около 2 тыс.). На рисунках 6 и 7 показаны фотографии их девушек и женщин в национальных одеждах. Это трудная для антропологов задача – выполнить их расовую идентификацию. Широкое лицо (в скулах) и высокая переносица до горбинки на носу. Небольшая уплощенность лица на уровне скул. Дополнительные трудности создает наличие на лице жировой прослойки, характерной для «степных» популяций. Тем не менее, мнение большинства антропологов совпадет: татарки не являются чистыми европеоидами. С другой стороны, идентифицировать их этнический компонент, как монголоидность (центральноазиатская) большинство антропологов не решится. Имеется в виду идентификация только по черепам. По внешнему же виду у большинства девушек и женщин явно просматриваются смягченные характеристики южносибирской расы.

Рис. 6 – Британский принц Чарльз среди татарских девушек, одетых в традиционные костюмы,
в деревне Кружиняна на севере Польши
Источник: https://birdinflight.com/ru/mir/neobychnye-etnicheskie-gruppy.html

Рис. 7 – Татарские женщины Литвы в национальных костюмах
Источник: https://baltnews.lt/vilnius_news/20170720/1017375845.html
10. Элементы реконструкций
10.1. Две базовые концепции этнических реконструкций
В рамках Традиционной истории историки, лингвисты, археологи и антропологи реконструировали прошлое венгров вглубь веков до 2000 г. до н. э. [Венгры]. Никаких проблем для интеграции в эту модель новых данных популяционной генетики не имеется. У венгров выявлены генетические маркеры, объединяющие их с хантами и манси, языки которых, как и венгерский входят в финно-угорскую группу. У мадьяр имеются восточноевразийские гаплогруппы и их субклады. Авары тоже легко локализуются в Южной или Восточной Сибири. По краниологическим данным автор публикации [Ery, 1983] локализовал прародины трех групп мадьяр в поздней фазе раннего железного века. Группа А – Северное Приаралье, группа В – нижнее течение Урала и Волги (без ее дельты), а также бассейн Дона (без его верхней части), группа С – бассейн Днепра (без его верхней части). Северное Приаралье рядом с югом Западной Сибири, региона проживания хантов и манси. То, что группы аваров, мадьяр и арпадов по краниологическим характеристикам почти неотличимы друг от друга, легко объясняется. Перед приходом на Среднедунайскую равнину они какое-то время проживали в одном и том же регионе, предположительно, на Южном Урале, в северном Приаралье и Северном Прикаспии.
Наши этнические реконструкции находятся в идеологическом поле Новой хронологии А.Т Фоменко и Г.В. Носовского. Основные ее элементы следующие [Носовский, Фоменко, 2012]. В XIV-XVI вв. в Евразии существовала Империя, созданная предками русских и популяций, говорящих сегодня на тюркских языках. Ее политический центр был в междуречье Волги и Оки. Армия базировалась в степной зоне Евразии и называлась Ордой. Военные компании по созданию Империи, колонизация славянами и тюрками малозаселённых территорий Европы, а также карательные экспедиции XIII-XV вв. в Традиционной истории «замаскированы» под внедрение в Центральную, Южную и Западную Европу славян, гуннов, аваров, готов, мадьяр и монголо-татар. Первые четыре фантома в ней фигурируют как элементы Великого переселения народов. Археологическое наследие реальных участников событий XIII-XV вв. – главным образом, дружинные погребения воинов Империи, археологи «привязали» к фантомам Традиционной истории. Их отнесли к скифам, сарматам, гуннам, готам, аварам, викингам, мадьярам, … Исходя из этого, территория Орды в степной и лесостепной зонах маркируется дружинными погребениями. На востоке она ограничена Тувой, Минусинской впадиной, Алтаем и прилегающими к нему территориями. На западе – Среднедунайской равниной.
10.2. Этнонимы
В тексте Анонима «Деяния венгров» («Gesta Hungarorum», латынь) относимом к 1196-1203 гг., сами мадьяры называют себя Scithici, а свою родину – Scithia. Конкретный регион, откуда они начали свой путь на запад – Dentumoger, а свое объединение – hetumoger [Шушарин, 1961]. Считается, что «hetu» в старовенгерском означает «семь», а слово hetumoger переводится как «семь мадьяр». Из этого ясно, что этноним «мадьяры» на латыни писался «moger». HETU – это вариант тюркского числительного «семь» – jete (башкирский), MOG – АЙМАК (АМ+АК, К<>Г, АК – суффикс), ER – суффикс. Это словосочетание тюркских слов означает «семь аймаков». В русском источнике XII в. «Повесть о латынех, когда отлучишасяот грек» имеется фраза «Пеони, глаголемее Оугри, иже сами нарицаются Магере...» (по [Шушарин, 1961]). То есть, современное самоназвание венгров magyarok (русское мадьяры) является специфической трансформацией их древнего этнонима moger/магере с суффиксом АК. В соответствии с правилами словообразования в тюркском языке moger примерно означает «аймачник». Так сама себя называла основная часть сообщества. Естественно, его элита аймачниками не являлась. Они были hungar или унгры, в русском – угры. Отметим, что в латинском варианте этнонима появился звук «Х» (по придыхательной схеме). Точно так же как он появился в тюркском слове jete. В русском выражении «куда Макар телят не гонял», Макар вовсе не имя, а магер. То есть, «туда, куда простой аймачник не гонял телят на выпас». В греческих свидетельствах мадьяр тоже назвали «пеоны». Это жители Паннонии (лат. Pannonia). Фразу из русской «Повести …» следует перевести так «Пенони (жители Паннонии), называемые уграми …». То есть, не все жители этого региона, но только конкретные угры.
В словосочетании DENTUMOGER (DEN+jTU+MOGER) ДЕН (уД+ИН, ИН – суффикс) – это то же самое, что и УДел. То есть, семь аймаков пришли из Скифии из удела семи аймаков. Регион семи аймаков по состоянию на вторую половину XVIII в. известен. Это владения башкирских родов объединения ете ырыу на юге Башкортостана и зона кочевок объединения казахских родов жетыру (еты = жеты) Младшего жуза казахов. Последняя локализована непосредственно южнее владений етеырыу. Ру и ырыу – «род». Отметим, что эти два объединения возникли в XVII-XVIII вв. «ниоткуда». Мы же предполагаем, что они являются реинкарнациями подразделения Орды «семь аймаков». По нашим лингвистическим реконструкциям оно было локализовано на юге Южного Урала и в Северном Приаралье.
Этнонимы сообществ, которые колонизировали Среднедунайскую равнину, хорошо известны. Это и тюркоязычные Kiskun и Nagykun. В последнем этнониме легко угадываются НОГАЙ. КУН в данном случае редуцированные суффиксы АК+АН, а вовсе не КУНы, в которых историки превратили КИМАКов. Последние – это тоже АЙМАКИ («К» по придыхательной схеме). ОНОГУРы письменных свидетельств, проживавшие на Северном Кавказе и в Приазовье, это тоже НОГАЙ с суффиксом АР. В данном случае не редуцирован первый гласный звук этого этнонима. Исходя из этого, этноним НОГАЙ (яН+аК+ИЙ, АК и ИЙ – суффиксы) восходит к ЯНЫ в тюркском «новый», ЯНГИ (ЯН+аК) в узбекском «новый», JUNG в немецком «молодой». В русском НОВЫЙ (яН+ОВ+ИЙ) и ЮННЫЙ (ЯН+иН+ИЙ). Ближайшая аналогия этнонима НОГАЙ – ЯНЫЧАР (ЯНы+ИЧ+АР, ИЧ и АР – суффиксы). Имеется аналогия ВЕНГР (ЯН+аК+аР, АК и АР – суффиксы) = HUNGARian. Здесь мы имеем две трансформации: У <> В и звук «Х», образовавшийся по придыхательной схеме. ЯНЫ в ДЖекающих тюркских диалектах звучит как ДЖАНЫ. Узбекское ЯНГИ будет звучать ДЖАНГИ. Это и есть CSANGO Трансильвании. Регион их проживания назывался Etelkoz. Он занимает важное место в реконструкциях завоевания мадьярами Среднедунайской равнины. ETELKOZ – это УДЕЛ KOZаков. На такой идентификации второй части названия региона не настаиваем. Выше мы показали только часть схемы трансформации звука «О». Ее полный вариант: О > ОУ > У > В > Б > П > Ф. То есть, имеется тождество ОНОГУР= UNGR = ВЕНГР = HUNGARian. Очевидная гипотеза UNGR = ОНОГУР приведена в справочнике [Венгры]. Наше дополнение к ней – это привязка венгров к ногаям Северного Кавказа.
По краниологическим данным мы сделали вывод: авары, мадьяры и арпады – это одно и то же сообщество. В прошлом это было известно. «Латиноязычные источники называли их [мадьяр] еще аварами или гуннами» [Пилипчук, 2015, с. 125]. С гуннами все ясно HUNgarian = HUNni (лат.) = ГУНны. То есть, основа этого этнонима тоже ЯНЫ. Страна ЯНов будет называться ПАННОНия (ЯН+АН+АН, АН – суффиксы). Все сошлось.
В этнониме KISKUN легко угадывается КАЗАК с суффиксом АН. Напомним, что предков казахов русские называли киргиз-КАЙСАКами и КАСАКами. Ираноязычные JASZ (YAZigian) – это ЯЗЫГи свидетельств, отнесенных к поздней Античности. Их активность на Дунае рассмотрена в публикации [Лысенко, 2006]. Если этноним ЯЗЫГ потеряет первый гласный звук, то это будет SZEKely (SEKlers). Последняя составляющая этого слова – ЭЛЬ, означает «сообщество», в тюркском ИЛ – «народ». В прошлом секлеров называли просто секеи [Поп, 2012]. Теперь можно перейти к основе этих этнонимов. Это лингвистический маркер АС/АЗ. Его общее значение – всякая верховная власть, земная и божественная [Тюрин, 2010, Асы]. Отсюда САР/ЦАРЬ (аС+АР, АР – суффикс), ИСА (Иисус), ЗАКОН (аЗ+АК+ИН) и ЯСА (АС+А). Воины, служащие закону (государству) – хАЗАРы (АЗ+АР) кАЗАКи (АЗ+АК) АСКЕРы (АС+аК+АР, АК и АР – суффиксы). Звуки «Х» и «К» в двух первых словах образовались по придыхательной схеме (Х > К). СКИФы (аС+аК+ОВ) – это сообщество, живущее по определенному закону. Люди закона. Или воины центральной власти. Вместе с Csango и секеями в Трансильвании проживали и проживают саксы. Они говорят на немецком языке. Считается, что это потомки колонистов из Саксонии и других немецких земель. Мы сформулируем другую гипотезу. Это такие же пришельца с востока, как и два других сообщества. А их немецкий язык связан с тем, что они в XVI в. приняли лютеранство. Понятно, что САКС – это то же самое, что СЕКЕЙ. Эти два слова отличаются суффиксами – ЕЙ и иШ. Главный вывод: этнонимы СКИФы, KISKUN, JASZ (YAZIGian), SZEKely (SEKlers) и САКСы – это варианты слова КАЗАК.
Csango, секеи и саксы жили в Transilvania. Это калька венгерского названия региона «Erde-elw (позже Erdely), то есть «Залесье» или «земля за лесом»» [Поп, 2012, с 56]. Мы же видим в словосочетании Erdely «ОРДынский ЭЛЬ».
«В самых ранних западноевропейских текстах, однако, их [мадьяр] называли turci или ungri» [Венгры]. Обращаем внимание на славянские формы двух слов – туркИ и унгрИ. Правители Венгрии АРПАДы – это немного трансформированное слово ОРДа (АР+УД, УД> ВУД > ПУД >ПАД). Это не имя, а титул. Точно так же титулом является АТТИЛА = УДЕЛ.
Вместе с мадьярами на Среднедунайскую равнину пришло три племени Kabars. Раньше они являлись частью сообщества хазар. Формы этого этнонима в разных свидетельствах рассмотрены в публикации [Toth, 2016]. Основные: Kabaroi, Qavars, Qabars, Khavars. Этноним сводится к АВАРЫ/ОБРЫ (АБ+АР, АР – суффикс), через «К» по придыхательной схеме. Для доказательства этой гипотезы нужно найти форму этого этнонима с начальным звуком «Х». Она имеется в справочнике [Венгерские]. Три племени, ушедшие от хазар названы «хабары».
Этническая группа венгров Paloc – это ПОЛЯК (ЭЛЬ+АК, АК – суффикс). То, что звук «П» образовался по придыхательной схеме, доказывает русская форма этого этнонима – ЛЯХ. То есть, ПОЛЯК и ПОЛОВЕЦ (ЭЛЬ+ОВ+ИЦ, ОВ и ИЦ – суффиксы) означает «член эля, член сообщества». Это то же самое, что и МАГЕР – простой аймачник. По нашей гипотезе этноним РУС (аР+ИШ, ИШ – суффикс) восходит к лингвистическому маркеру АР. В тюркском этот этноним звучит как УРУС. Это и есть ORSEG (АР+иШ+АК), то есть, РУССКий. Но, «its mediaeval form was still urusag» [Endrey, 1975, p. 79]. Этнониму urusag соответствует этноним тувинского рода ооржак.
10.3. Популяционная генетика
Регион формирования сообществ, которые колонизировали Среднедунайскую равнину, хорошо известен. В первом приближении это улус Джучи – Северное Причерноморье, Северный Кавказ, Нижнее и Среднее Поволжье, Южный Урал, регион Арала, юг Западной Сибири. В соответствии с нашей трактовкой археологических культур, к этому региону нужно отнести и Южную Сибирь, прежде всего, зону скифской культуры, которая включает Алтай, Минусинскую впадину и территорию Тувы. В обозначенном регионе основная часть погребений, относимых археологами к бронзовому и раннему железному векам, являются дружинными. В них погребены воины Орды. Поэтому мадьяры близки к представителям археологических культур по ассоциациям генетических маркеров и краниологическим характеристикам. Близки они и к популяциям региона – татарам, башкирам и тувинцам. Авары по гаплогруппам Y-хромосомы близки к тувинцам.
Позднее, в регион базирования Орды была включена и Среднедунайская равнина. Туда было переведено несколько орд, на основе которых сформировались этнические группы венгров Kiskun, Nagykun, Jasz, Csango, секлеры. Ираноязычные Jasz вполне могли быть из Приаралья. Часть популяций Хорезмского оазиса до прихода туда кочевых узбеков была ираноязычной. В новом регионе базирования подразделений Орды был осуществлен призыв в них населения сопредельных областей, носителей субкладов гаплогрупп R1a, R1b, I2 и E. Суммарно их частоты у мадьяр 63,3 %. Они явились в Орду без своих женщин, носителей заподноевразийских гаплогрупп. Поэтому в жены брали женщин рожденных в Орде. А у них была относительно высокая доля восточноевразийских гаплогрупп. Этим объясняется высокие частоты последних у мадьяр. Авары же пришли в восточную Европу с «неполным комплектом» своих женщин, носителей восточноевразийских гаплогрупп. Им пришлось брать в жены западных славянок. На диаграмме главных компонент полногеномных маркеров (Рис. 2) Орде соответствует кластер III.
Литовско-татарский парадокс разрешается просто. Литовские татары, как субэтнос сформированы из потомственной военной элиты постордынских политических образований. В формировании ордынской элиты в XIII-XIV вв. представители популяций Южной Сибири не участвовали. Поэтом их потомков по мужской линии среди литовских татар не имеется. Женщины же татар в первом приближении характеризуют «степной» контингент, по генетическим характеристикам близкий к южносибирской расе.
10.4. Венгерский язык
В рамках Новой хронологии достигнуто понимание двух реалий:
-большинство письменных языков является искусственными, причем, при их формировании решались политические задачи;
- современная лингвистика имеет договорные основы.
Во второй половине XIX в. в Европе началось формирование этносов. Этот процесс управлялся региональной элитой. Ее группы формировали «свои» этносы. Одним из главных элементов процесса было создание искусственных письменных языков. На территории Австрийской империи государственным языком являлся немецкий. Широко использовалась латынь. Элита говорила и на французском языке. В этих условиях региональная элита Среднедунайской равнины начала формировать венгерский этнос.
«Современный венгерский языквозник только в XVIII в.» [Пилипчук, 2015, с. 125]. Это не совсем так. В конце XVIII в. началось его формирование. Но жесткая коллизия, называемая «Угро-тюркским противостоянием» случилась только во второй половине XIXв. «В спорах о генезисе венгерского народа и его языка, о взаимосвязи общих корней с финно-угорскими, тюркскими и монгольским языками принимали активное участие […] выдающиеся ученые-востоковеды Европы – Арминий Вамбери (1832-1913 гг.), венгерский востоковед, путешественник, полиглот, и Йозеф Буденц (1836-1892 гг.), немецкий языковед, профессор Будпештского университета, член-корреспондент Санкт-Петербургской академии наук» [Валеев, 2018, с. 29]. Первый считал, что венгерский язык имеет тюркскую основу, второй – финно-угорскую. Габор Балинт (1844-1913 гг.) – выдающийся венгерский языковед, монголовед и тюрколог,обосновал третью точку зрения: «венгерский язык является отдельной ветвью, а не частью финно-угорской или тюркской (алтайской) языковой семьи» (с. 30).
В академических дискуссиях обсуждались вовсе не этнические корни венгерского языка. Вопрос был в другом. Каким будет искусственный письменный венгерский язык. По каким-то соображениям решили (не лингвисты) что он будет относиться к финно-угорской группе уральской языковой семьи. Под это лингвисты сформировали его словарный базис из различных диалектов этнических групп венгров, а элита начала внедрять новодел в массы. И сегодня среди лингвистов действует договор – сомнений в правильности идентификации венгерского языка не имеется. Под нее археологами «подтянуты» соответствующие материальные основания. Все закреплено намертво.
В общеупотребительной венгерской лексике непроизводные слова финно-угорского происхождения составляют 10,1%, славянизмы 9,36%, германизмы5,43 %, тюркизмы 4,59% [Золтан, 2013]. При этом, исходя из вышесказанного, многие слова первого пласта в язык имплантированы искусственно. То есть, доля слов финно-угорского происхождения в венгерском языке по его состоянию на период «Угро-тюркского противостояния» не превышала долю славянизмов. По особенностям славянских слов в венгерском языке установлено, что они попали в него из диалекта болгарского типа. Но здесь у лингвистов возможны аберрации. Считается, что церковно-славянский язык создан на основе древнеболгарского. То есть, славянизмы в венгерском языке могут быть из церковно-славянского языка, а он являлся общеславянским. В публикации [Пилипчук, 2015] отметается, что «венгры переняли у славян [западных и южных] военную лексику» (с. 124). Это в высшей степени странно. Завоеватели перенимают у завоеванных военные термины. По нашему мнению, военная лексика венгров является общеславянской и попала в их язык от элиты Орды, основа которой была русской. Такой состав венгерского языка полностью соответствует данным популяционной генетики и краниологии, а также нашим этническим реконструкциям. Он является миксом славянского, финно-угорского, немецкого и тюркского языков. Имеются в нем и иранский массив слов.
Но не все лингвисты придерживаются отмеченного выше договора. В публикации [Валиева, 2014] рассмотрены булгаризмы башкирского и венгерского языков. Вывод: «в основе обоих языков лежит общаятюркская лексика» (с. 155). С другой стороны, имеется большое количество публикаций, в которых отражены общие элементы венгерского языка и языков уральской семьи. То есть, «Угро-тюркское противостояние» продолжается. В заключение приведем свидетельство филолога узбечки. Находясь в Венгрии, она понимала целые блоки устной речи.
11. Общие результаты
1. По данным популяционной генетики венгры не отличаются от соседних с ними популяций. Но у них имеются линии гаплогруппы Y-хромосомы N (1,2 %), характерные для хантов и манси, а также популяций Южного Урала, Волго-Уральского региона и, возможно, Южной Сибири.
2. По частотам гаплогрупп Y-хромосомы и их субкладам мадьяры похожи на поволжских татар, авары – на тувинцев. По частотам гаплогрупп мтДНК мадьяры близки европейским скифам, представителям потаповской и полтавкинской археологических культур, а также поволжским татарам. Их западноевразийские гапогруппы близки к гаплогруппам представителей срубной культуры, а восточноевразийские – тувинцев.
3. По краниологическим характеристикам авары, мадьяры и арпады являются одним сообществом, в котором выделяется четыре субпопуляции. Одна соответствует восточным и западным славянам. Другая имеет характеристики близкие к южносибирской расе. Две субпопуляции – европеоиды с элементами монголоидности и уралоидности.
4. Этнические реконструкции выполнены в идеологическом поле Новой хронологии А.Т Фоменко и Г.В. Носовского. Единое сообщество, пришедшее врегион в XIII-XIV вв. (в Традиционной истории авары и венгры), сформировано в Орде, армии Империи. Территория базирования Орды включала степную и лесостепную зоны Евразии от Северного Причерноморья до Минусинской впадины и Тувы. Воины ее отдельных подразделенийи их жены имели значимый востчноевразийский генетический компонент. Этим этническим реконструкциям соответствует словарная база венгерского языка. Он является миксом славянского, финно-угорского, немецкого, тюркского и иранского языков.
Литература
Акимова М.С. Некоторые итоги изучения антропологического состава древнего населения Прикамья // Вопросы археологии Урала, 1961, Вып. 1, с. 121-129.
Валеев Р.М., Валеева Р.З., Мартынов Д.Е., Сахавова М. Венгерский монголовед БалинтГабор в Казани: Неизвестные письма Н. И. Ильминскому (1872-1874 гг.) // Первые Ковалевские чтения, 2018, с. 28-31.
Валиева М.Р. Булгаризмы в апеллятивной лексике башкирского и мадьярского (венгерского) языков // Вестник ВЭГУ, 2014, № 3 (71), с. 155-160.
Венгры. Происхождение и ранняя история. Всемирная история. http://www.world-history.ru/nations_about/1839.html
Венгерские литература и язык. Энциклопедический Словарь Ф.А. Брокгауза и И.А. Ефрона. В 86 полутомах с иллюстрациями и дополнительными материалами, С.-Петербург, 1890-1907.
Герасимов М.М. Основы восстановления лица по черепу. М. Советская наука, 1949, 188 с.
Герасимова М.М., Рудь Н.М., Яблонский Л.Т. Антропология античного и средневекового населения Восточной Европы, 1987, 253 с.
Дамба Л.Д., Балановская Е.В., Жабагин М.К., Юсупов Ю.М., Богунов Ю.В., Сабитов Ж.М., Агджоян А.Т., Короткова Н.А., Лавряшина М.Б., Монгуш Б.Б., Кавайоол У.Н., Балановский О.П. Оценка вклада монгольской экспансии в генофонд тувинцев // Вавиловский журнал генетики и селекции, 2018, Т. 22, № 5, с. 611-619.
Деренко М.В., Дамбуев И.Л, Малярчук Б.А. и др. Структура и разнообразие митохондриального генофонда коренного населения Тувы и Бурятии по данным о рестрикционном полиморфизме // Генетика, 1999, Т. 35, № 12, с. 1706-1712.
Золтан A. (Zoltan A.) Славянские диалекты Карпатского бассейна во время прихода венгров (IXв.) Studia Slavica Hung. 58/1 209-218. (2013).
Исмагулов О., Исмагулова А.О., Надирбеков И.О., Сатаев М.А. Динамика краниологических показателей древних и средневековых насельников Казахстана в свете этногенеза казахского народа // Известия Самарского научного центра Российской академии наук. Социальные, гуманитарные, медико-биологические науки, 2018, Т. 20, № 5, с. 59-72.
Кириченко Д.А. Сарматы Венгрии по данным антропологии // Вестник археологии, антропологии и этнографии, 2013, № 3 (22), с. 103-112.
Клёсов А.А. Секлеры восточной Европы и их гаплогруппы и гаплотипы // Вестник Российской Академии ДНК-генеалогии, 2009, Том 2, № 1, с. 2-51.
Клёсов А.А. Венеты и венеды по данным ДНК-генеалогии // Исторический Формат, 2015, № 2, с. 75-102.
Козинцев А.Г. Рецензия на книгу «Антропологические данные к вопросу о великом переселении народов. Авары и сарматы (Т.А. Тот, Б.В. Фирштейн, 1970 г.) // Советская этнография, 1972, № 6, с.165-167.
Конюхова Е.В., Нгуен Фыонг Нга, Алимова Ф.К., Кравцова О.А. Разнообразие митохондриального генофонда в популяции поволжских татар Республики Татарстан // Ученые записки Казанского государственного университета, 2010, Том 152, кн. 2, с. 128-136.
Лобов А.С. Структура генофонда субпопуляций башкир. Автореферат диссертации на соискание ученой степени кандидата биологических наук. Уфа, 2009, 23 с.
Лысенко Н.Н. Языги на дунайском лимесе Рима в I-II вв. н.э. // Нижневолжский археологический вестник, 2006, Вып. 8, с. 139-153.
Матюшко И.В. Особенности погребального обряда кочевников степного Приуралья XIII-XIV вв. // Известия Самарского научного центра Российской академии наук, 2011, Т. 13, № 3-1, с. 280-283.
Носовский Г.В., Фоменко А.Т. Империя. Славянское завоевание мира. Европа. Китай. Япония. Русь как средневековая метрополия Великой Империи. АСТ, 2012, 830 с.
Панкратов В.С., Кушнеревич Е.И., Чеботарев Л.Ю., Мецпалу Э., Давыденко О.Г. Формирование пула митохондриальной ДНК белорусских татар: дальние миграции и смешение генофондов. // Доклады Национальной академии наук Беларуси, 2014, Т. 58, № 3, с. 82-87.
Пилипчук Я.В. Предыстория венгров и венгеро-пермская проблема // Гасырларавазы, 2015, № 1-2, с. 121-132.
Поп И.А. Генезис средневековой государственности в Трансильвании (IX-XIII вв.) // Studia Slavicaet Balcanica Petropolitana, 2012, № 1 (11), с. 51-78.
Рожанский И.Л. Литовские татары. ДНК-родословные и их корни в степях Евразии // Исторический Формат, 2016, № 4, с. 89-105.
Руденко К.А., Нечвалода А.И. Могильник на Тетюшском II городище в Татарстане: портретная антропологическая реконструкция по черепу и краниология одного погребения // Теория и практика археологических исследований, 2016, № 3 (15), с. 126-149.
Тот Т.А. Древнейшие периоды в этногенезе венгерского народа // Вопросы иранской и общей филологии, 1977, с. 280-291
Трепавлов В.В. История Ногайской Орды. 2-е изд., испр. и доп. Казань: Издательский дом «Казанская недвижимость», 2016, 764 с
[Тюрин, 2010, Генетические следы] Тюрин А.М. Имеются ли генетические следы монгольских завоеваний 13 века? // Электронный сборник статей «Новая Хронология». Вып. 10, 2010. [Новая хронология]
[Тюрин, 2010, Венгры] Тюрин А.М. Согласование исторических свидетельств, лингвистических и генетических данных, характеризующих венгров // Электронный сборник статей «Новая Хронология», 2010, Вып. 10. [Новая хронология].
[Тюрин, 2010, Асы] Тюрин А.М. Асы, ясы, языги, осетины, казаки, казахи, гузы, усюны и узбеки. Полемика, 2010. [Новая хронология]
[Тюрин, 2017, Ногаи, генетика] Тюрин А.М. Калмыки, караногайцы, кубанские ногайцы и крымские татары – геногеографический и геногенеалогический аспекты // Журнал фронтирных исследований, 2017, № 2, с. 7-29.
[Тюрин, 2017, Литовские татары] Тюрин А.М. Генетический портрет литовских татар и феномен «Монгольские завоевания 13 века» // Вестник Оренбургского государственного университета, 2017, № 5, с. 78-82.
[Тюрин, 2017, Казахские торе] Тюрин А.М. Казахские торе не являются потомками Чингисхана // Цивилизация знаний: российские реалии: труды Восемнадцатой Международной научной конференции, Москва, 21–22 апреля 2017 г. – М.: РосНОУ, 2017, с. 93-94.
[Тюрин, 2018, Ногаи, антропология] Тюрин А.М. Ногаи: антропологический аспект // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2018, Монголоидность, сарматы] Тюрин А.М. Датирование сарматов по монголоидности // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2019, Караногайцы] Тюрин А.М.Элементы этногенеза караногайцев по данным популяционной генетики // Перекрёстки истории. Актуальные проблемы исторической науки, 2019, с. 56-60.
[Тюрин, 2019, Кубанские ногайцы] Тюрин А.М. Элементы этногенеза кубанских ногайцев по данным популяционной генетики // Перекрёстки истории. Актуальные проблемы исторической науки, 2019, с. 60-64.
[Тюрин, 2019, Башкиры-1] Тюрин А.М.К вопросу о монголоидных компонентах у башкир // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2019, Башкиры-2] Тюрин А.М. К вопросу о монголоидных компонентах у башкир // XIII Конгресс антропологов и этнологов России: сб. материалов. Казань, 2–6 июля 2019 г. / Отв. ред.: М.Ю. Мартынова. – Москва; Казань: ИЭА РАН, КФУ, Институт истории им. Ш. Марджани АН РТ, 2019, с. 95.
[Тюрин, 2019, Викинги] Тюрин А.М.Эра викингов, Норманнская теория, популяционная генетика и Новая хронология. Препринт, 2019. [Новая хронология].
[Тюрин, 2019, Скифы] Тюрин А.М. Популяционная генетика: расовая и этническая идентификация киммерийцев, скифов, сарматов, саков и черняховцев. Препринт, 2019. [Новая хронология].
Харьков В.Н., Хамина К.В., Медведева О.Ф., Симонова К.В., Хитринская И.Ю., Степанов В.А. Структура генофонда тувинцев по маркерам Y-хромосомы // Генетика, 2013, Т. 49, № 12, с. 1416.
Чикишева Т.А. Антропология носителей пазырыкской культуры // Феномен алтайских мумий, Новосибирск, 2000 (Гл.1.3), с. 35-49.
Шушарин В.П. Русско-венгерские отношения в IX веке // Международные связи России до XVII века, 1961, с. 131-180.
Balinova, N. et al. Y-chromosomal analysis of clan structure of Kalmyks, the only European Mongol people, and their relationship to Oirat-Mongols of Inner Asia. Am. J. Hum. Genet.27, 1466-1474 (2019).
Battaglia, V. et al. Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe. Eur. J. Hum. Genet. 17(6):820–30 (2009).
Brandstatter, A. et al. Migration rates and genetic structure of two Hungarian ethnic groups in Transylvania, Romania. Ann. Human Genetics, 71, 791-803 (2007).
Cocos, R.Genetic affinities among the historical provinces of Romania and Central Europe as revealed by an mtDNA analysis. BMC Genetics, vol. 18, Article number: 20 (2017).
Csanyi, B. et al. Y-chromosome analysis of ancient Hungarian and two modern Hungarian-speaking populations from the Carpathian Basin. Ann. Hum. Genet. 72, 519-534 (2008).
Csosz, A. et al. Maternal Genetic Ancestry and Legacy of 10th Century AD Hungarians. Sci. Rep. 6, 33446 (2016).
Endrey, A. Sons of Nimrod: the origin of Hungarians. Melbourne: The Hawthorn Press, 1975, 116 p.
Ery, K. Comparative statistical studies on the physical anthropology of the Carpathian Basin populations between the 6–12th centuries A.D. Alba Regia 20, 89-141 (1983).
Family Tree DNA Hungarian_Magyar_Y-DNA_Project. Familytreedna.com. Retrieved 18 August 2017. https://www.familytreedna.com/public/Hungarian_Magyar_Y-DNA_Project?iframe=yresults
Guglielmino, C.R., Judit, B. Genetic structure in relation to the history of Hungarian ethnic groups. Hum. Biol. 68(3):335-55 (1996).
Guglielmino, C.R. et al. Probable ancestors of Hungarian ethnic groups: an admixture analysis. Ann Hum Genet. 64(Pt 2):145-59 (2000).
Haplogroup R1a. http://blog.vayda.pl/en/haplogroup-r1a-statistic-02-2018-14-new/
Järve, M. et al. Shifts in the Genetic Landscape of the Western Eurasian Steppe Associated with the Beginning and End of the Scythian Dominance // Current Biology, Vol. 29, Issue 14, рp. 2430-2441 (2019)
Juras, A. et al. Diverse origin of mitochondrial lineages in Iron Age Black Sea Scythians. Sci. Rep. 7, 43950 (2017).
Malyarchuk, B.A. et al. Mitochondrial DNA variability in the Czech population, with application to the ethnic history of Slavs. Hum Biol 78: 681-696 (2006).
Maroti, Z. et al. MITOMIX, an Algorithm to Reconstruct Population Admixture Histories Indicates Ancient European Ancestry of Modern Hungarians. biRxiv. Preprint (2018).
Movsesian, A.A., Bakholdina V. Nonmetric cranial trait variation and the origins of the Scythians, American Journal of Physical Anthropology. 162 (3) (2017).
Naumova, O.Yu., RychkovS.Yu., Zhukova O.V. Mitochondrial DNA Variability in Populations and Ethnic Groups of Tatars of the Tobol–Irtysh Basin. Genetika.; 45(9):1260-9 (2009).
Neparaczki, E. et al. Mitogenomic data indicate admixture components of Central-Inner Asian and Srubnaya origin in the conquering Hungarians. PLoS ONE 13(10): e0205920 (2018).
Neparaczki, E. et al. Y-chromosome haplogroups from Hun, Avar and conquering Hungarian period nomadic people of the Carpathian Basin. Preprint. (2019).
Olasz, J. et al. DNA profiling of Hungarian King Béla III and other skeletal remains originating from the Royal Basilica of Székesfehérvár. Archaeological and Anthropological Sciences, Vol. 11, Issue 4, pp 1345-1357 (2019).
Narasimhan, V.M. et al. The formation of human populations in South and Central Asia. Science, Vol. 365, Issue 6457, eaat7487 (2019).
Olasz, J. et al. DNA profiling of Hungarian King Béla III and other skeletal remains originating from the Royal Basilica of Székesfehérvár. Archaeological and Anthropological Sciences, Volume 11, Issue 4, pp 1345-1357 (2019).
Pamjav, H. et al. A study of the Bodrogkцz population in north-eastern Hungary by Y chromosomal haplotypes and haplogroups. Mol. Genet. Genomics 292, 883–894 (2017).
Pankratov V. et al. East Eurasian ancestry in the middle of Europe: genetic footprints of Steppe nomads in the genomes of Belarusian Lipka Tatars. Sci Rep. 25;6:30197 (2016).
Post, H. et al. Y-chromosomal connection between Hungarians and geographically distant populations of the Ural Mountain region and West Siberia. Scientific Reports, vol. 9, 7786 (2019).
Semino, O. et al. MtDNA and Y chromosome polymorphisms in Hungary: Inferences from the palaeolithic, neolithic and Uralic influences on the modern Hungarian gene pool. Eur. J. Hum. Genet. 8, 339-346 (2000).
Tambets, K. et al. The western and eastern roots of the Saami – The story of genetic «outliers» told by mitochondrial DNA and Y chromosomes. Am. J. Hum. Genet. 74, 661-682 (2004).
Toth, S.L. The Qavars (Qabars) and their Role in the Hungarian Tribal Federation. Chronica: Annual of the Institute of History, University of Szeged, 12 3-22. (2016).
Volgyi, A. et al. Haplogroup distribution of Hungarian population and the largest minority group. Forensic Science International: Genetics Supplement Series.
Vol. 1, Issue 1, 383-385 (2008).
Yunusbayev, B. et al. Thegenetic legacy of the expansion of Turkic-speaking nomads across Eurasia // PLoSGenet. V.11(4):e1005068 (2015).
статья получена 23.10.2019
