Согласование исторических свидетельств, лингвистических и генетических данных, характеризующих венгров
Тюрин Анатолий Матвеевич, к.г.-м.н.
Ключевые слова: Новая Хронология, венгры, история, генетика, лингвистика.
Оглавление
1. Формальная постановка задачи
2. Исторический аспект
3. Генетический аспект
5. Генетические данные по древним жителям Южной Сибири
6. Реконструкции
Источники информации
1. Формальная постановка задачи
По крайней мере, три кочевых народа, фигурирующих в Традиционной Истории (ТИ), имеют общие характерные элементы своих деяний. Это гунны, авары и мадьяры. Они сформировались в Центральной Азии, активно воевали в Европе, а затем поселились на Среднедунайской равнине. Имеются ли следы этих пришельцев в современном генетическом портрете венгров, жителей Среднедунайской равнины? Поиск ответа на этот вопрос – задача минимум. Задача максимум – согласование исторических свидетельств о предках венгров и новейших генетических данных. При этом согласовании приняты во внимание и лингвистические данные. Таким образом, постановка задачи соответствует названию статьи.
2. Исторический аспект
Собственно история венгров началась в 9 веке, когда мадьяры поселились на Среднедунайской равнине. Они говорили на том же языке, на котором говорят современные венгры. Он относится к финно-угорской группе уральской языковой семьи. До прихода на Среднедунайскую равнину мадьяры жили в центрально-азиатских степях.
Среди современных венгров выделяется 8 этнических групп [Guglielmino, 2000]. Группа Orseg живет около юго-западной границы Венгрии (рисунок 1) в условиях генетической изоляции со времени прихода мадьяр. Группа Paloc живет, по крайней мере, со Средних веков на севере Венгрии и говорит на специфическом венгерском диалекте. Регион их проживания был оккупирован аварами в 6 веке. Группа Matyo живет восточнее Paloc. В свидетельствах 1275 года регион упоминался как «необитаемый», но около 1500 года он был заселен жителями сопредельных областей. Сегодня Matyo является изолированной группой. Это, возможно, связано с тем, что они католики, а жители соседних областей – протестанты. Группа Szekely (Seklers) живет в Трансильвании (Румыния) в Буковине. Они считают себя венграми, потомками Гуннов (Huns). В 11-12 веках Szekely поселились в регионе, который называется сегодня Szekely Land. Они отразили атаки печенегов (Pecheneng) и куманов (Kun) поскольку их мужчины были воинами (they were mainly soldiers). Группа Csango (Chango) живет в Молдавии в регионе, который назывался Etelkoz. Это одна из групп мадьяр, осевших в этом регионе во время завоевания ими в 896 году Венгрии. По другой гипотезе они поселились здесь после монгольских завоеваний 13 века. Группы Kiskun и Nagykun (их еще называют Small and Great Cumanians (Kuns)) считают своих предков тюрками, которые пришли в Венгрию во время монгольского завоевания 1241-42 годов. Эти две группы разделены Тисой. Группа Jasz (Yazigian) пришла в Венгрию вместе с куманами (Kuns) во время монгольского вторжения. Она имеет иранское происхождение и до 16 века говорила на иранском языке «used an Iranian language». В публикации [Guglielmino, 1996] приводятся некоторые уточняющие сведения по этническим группам венгров. Предки Kiskun и Nagykun до прихода в Венгрию жили в западной части Центральной Азии и говорили на тюркском языке. Кальвинизм (христианство протестантского толка) они приняли в 16 веке. Jasz пришли в Венгрию после татарского вторжения («They arrived in Hungary after the Tatar invasion (1241-1242)»). Они приняли католическую форму христианства.
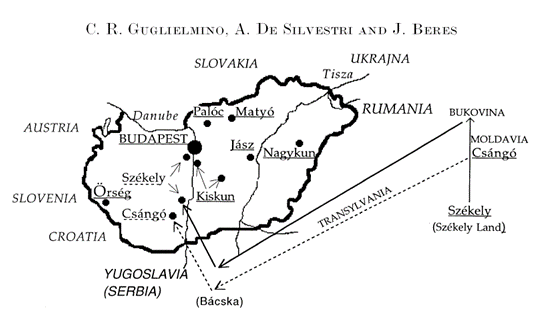
Рисунок 1. Этнические группы венгров [Guglielmino, 2000].
Стрелками показана миграция части венгров Szekely и Csango во время и после Второй Мировой войны.
3. Генетический аспект
3.1. Общие замечания
В последние годы форсированными темпами развиваются генетические исследования Человека. Применительно к изучению его прошлого обособились три дисциплины: геногенеалогия, геногеография и генохронология, которые являются составными частями генетики. Они базируются на одном и том же массиве данных – генетических маркерах человека.
Различают два типа генетических маркеров. Генетические маркеры (гены), которые:
- не влияют на биологические особенности людей, то есть, не определяют их «конкурентоспособность» в выживании в определенных природных и социальных условиях;
- определяют биологические особенности людей и являются объектом естественного отбора.
Имеются маркеры, не влияющие на биологические особенности людей, которые передаются только от отца к сыну (их выделяют в Y-хромосоме) или от матери к дочери (их выделяют в мтДНК). В Y-хромосоме выделяются два вида маркеров. Они определяют «гаплогруппу» и «гаплотип» людей. Но гаплогруппа не является совокупностью гаплотипов. Это самостоятельный маркер. В маркерах мтДНК тоже выделяют гаплогруппы и гаплотипы. Ко второму виду генетических маркеров относятся, например, гены, которые ответственны за группу крови человека и ее резус-фактор.
3.2. Конкретная постановка задачи
По решаемой нами задаче, относительно достоверно известно следующее:
- автохтонное население Сренедунайской равнины до прихода на нее переселенцев с востока не отличалось от жителей Центральной Европы и говорило на европейских языках;
- с востока на Сренедунайскую равнину пришли и на ней остались мадьяры, гунны, куманы и другие социальные общности;
- те, кто пришел и остался, говорили на венгерском, тюркском и иранском языках;
- с территорией Венгрии связана деятельность авар, монголо-татар и печенегов.
Менее достоверно известно о том, что предки социальных общностей, которые мы перечислили выше, жили в Центральной Азии и не являлись народами, родственными другим народам Центральной Европы.
На основе этих исторических данных конкретная исследовательская задача формируется предельно просто. Необходимо выявить среди венгров специфические генетические маркеры, свидетельствующие о том, что часть их предков была родственна народам Центральной Европы, а другая часть – народам Центральной Азии и Сибири. Именно так формулировалась задача, при изучении генофонда венгров. Но это ущербная постановка задачи, поскольку она базируется на принципе актуализма, заданном в неявном виде. Корректная постановка задачи звучит так: создание генетического портрета социальных общностей, пришедших в Венгрию с востока. Решается эта задача тоже предельно просто. Среди современных венгров необходимо выявить генетические маркеры, не характерные для жителей Центральной Европы. После того, как эта задача будет решена, можно попытаться выполнить геногенеалогические и геногеографические реконструкции, то есть определить регионы, откуда могли придти на территорию Венгрии социальные общности, имеющие конкретные характеристики генетического портрета.
3.3. Общегенетические характеристики венгров
Первые попытки поиска предков венгров по генетическим данным были предприняты в 90-х годах 20 века [Guglielmino, 1996, 2000; Pap, 2000]. Анализировались генетические маркеры, которые определяют биологические особенности людей. Внимания заслуживают только результаты обработки данных по 8 этническим группам венгров [Guglielmino, 2000], которые рассмотрены в разделе 2. По ним было сформировано 8 выборок генетических данных. Было сформировано еще несколько выборок. Три выборки характеризуют венгров Будапешта, а также юго-запада и севера Венгрии. Это так называемые смешанные выборки венгров. Здесь важно понимать, что выборки по этническим группам характеризуют именно тех венгров, предки которых пришли с востока, а смешенные выборки характеризуют всех венгров, в том числе и тех, предки которых являлись автохтонными жителями Среднедунайской равнины. Еще 4 выборки характеризуют европейские народы – итальянцев, немцев, поляков, а так же славян (Slavs). Последняя выборка включает сербов, болгар, чехов и русских. Другие 6 выборок характеризуют народы, предки которых могли быть родственны предкам венгров, пришедших с востока. Это турки, финны, иранцы, саамы (Lapps), а также уральские (Uralics) и восточные (Orientals) народы. Первые включают самодийцев, ненцев, энцев, нганасан, коми и мари-черемисов, вторые – монголов.
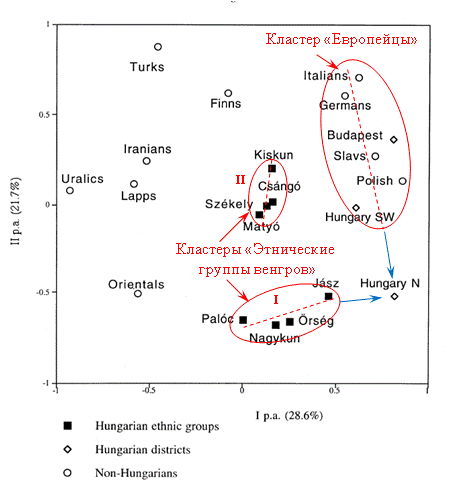
Рисунок 2. Положение точек, характеризующих 21 популяцию, в поле главных компонент [Guglielmino, 2000].
Красным и синим цветами показана информация, нанесенная А.М. Тюриным. Пояснения даны в тексте статьи.
В кластер «Этнические группы венгров» I попали точки «Nagykun» (характеризует группу, говорившую на тюркском языке), «Paloc» и «Orseg», (характеризуют группы, говорившие на венгерском зыке) и «Jasz», (характеризует группу, говорившую на иранском языке). Причем, точка «Nagykun» находится между точками «Paloc» и «Orseg». Три точки кластера – «Paloc» «Jasz» и «Nagykun», характеризуют группы венгров, живущих в близко расположенных районах Венгрии (рисунок 1). Другая точка кластера – «Orseg», характеризует группу, район проживания которой находится «в другом конце» Венгрии. В кластер «Этнические группы венгров» II попали точки «Szekely», «Csango», «Matyo» и «Kiskun». Последняя точка характеризует группу, говорившую на тюркском языке. То есть две точки, характеризующие этнические группы венгров, говоривших на тюркском языке – «Kiskun» и «Nagykun», попали в разные кластеры. Точки «Szekely» (венгры Буковины) и «Csango» (венгры Молдавии) практически наложились друг на друга. В публикации [Brandstatter, 2007] приведены результаты анализа мтДНК 360 человек из популяций венгров Szekely и Csango. Изучались и гаплотипы людей. Установлено, что генный обмен между популяциями был низким. Популяция Csаngо находилась в генетической изоляции. Сделан вывод о том, что предки популяций до прихода в Трансильванию жили в близко расположенных регионах.
По рассмотренным генетическим данным и результатам их интерпретации можно сделать следующие выводы.
1. Венгры являются типичной европейской нацией при некоторых специфических особенностях жителей севера Венгрии. Причем эти особенности связаны с этническими группами венгров Paloc, Jasz и Nagykun.
2. Все этнические группы венгров отличаются от европейцев. Но их близость к каким-либо народам Евразии не просматривается.
3. Этнические группы венгров четко разделились на два кластера. Возможно, это связано с тем, что они имели две разные предковые популяции, близкие по своим генетическим характеристикам.
4. Этнические группы венгров Paloc, Orseg, Kiskun и Jasz, предки которых говорили на венгерском, тюркском и иранском языках генетически не различаются. Это означает, что их предки входили в одно и то же социальное сообщество, части которого пришли в Венгрию возможно разными путями и в разное время. Это же относится и к группам Szekely, Csango, Kiskun и Matyo.
3.4. Гаплогруппы Y-хромосомы
Для решения поставленной задачи можно ограничиться рассмотрением гаплогрупп N3, C и Q. Гаплогруппа N3 присутствует практически у всех социальных общностей, говорящих на финно-угорских языках, а так же у их соседей. Гаплогруппа Q характерна для некоренных сибирских народов, гаплогруппа C – для монголов. Но одна из этнических групп венгров – Jasz, говорила на иранском языке. Если предки Jasz пришли с территории Ирана, то мы не сможем выявить их потомков среди современных венгров по маркерам, обозначенным выше. Учитывая близость компонентного состав гаплогрупп народов Ирана к европейскому, искать иранские корни Jasz можно только на основе «тонких» структур Y-хромосом и мтДНК. Выполнение этой работы находится за пределами наших возможностей.
В таблице 1 приведены частоты ключевых гаплогрупп Y-хромосомы народов Евразии [Tambets, 2004]. В Урало-Поволжском регионе максимальные частоты гаплогруппы N3 выявлены у удмуртов – 56,3%. У марийцев частота этого маркера – 31,5%, мордвы – 16,9%, коми – 22,3%. Эти народы говорят на языках финно-угорской группы. Частоты гаплогруппы N3 у татар и чувашей – 18,3% и 17,7%, соответственно. У других народов, говорящих на финно-угорских языках, выявлены следующие частоты гаплогруппы N3: саамы – 47,2%, финны – 63,2%, эстонцы – 30,6%, ханты – 38,3%.
Частоты ключевых гаплогрупп Y-хромосомы народов Евразии [Tambets, 2004]
(из таблицы исключены пояснения и ссылки на источники информации).
| Region and Population | Language Family | Sample Size |
Frequency of Haplogroup (%) | ||||||
| I | R1b | R1a | N3 | N2 | Q | C | |||
| Scandinavia | |||||||||
| Saami | U-FU | 127 (58) | 25,9 | 3,9 | 11,0 | 47,2 | 0 | 0 | 0 |
| Swedes | IE-G | 141 | 48,2 | 22,0 | 18,4 | 2,8 | 0 | 0 | 0 |
| Norwegians | IE-G | 72 | 40,3 | 27,8 | 23,6 | 6,9 | 0 | 0 | 0 |
| Danes | IE-G | 194 | 38,7 | 36,1 | 16,5 | 0,5 | 0 | 0 | 0 |
| Northern Europe | |||||||||
| Finns | U-FU | 38 | 28,9 | 0 | 7,9 | 63,2 | 0 | 0 | 0 |
| Estonians | U-FU | 209 | 18,2 | 9,1 | 33,5 | 30,6 | 0 | 0 | 0 |
| Latvians | IE-B | 86 | 7,0 | 9,3 | 38,4 | 41,9 | 0 | 0 | 0 |
| North Russians | IE-S | 77 | 20,8 | 0 | 40,3 | 28,6 | 0 | 0 | 0 |
| Eastern Europe | |||||||||
| Russians | IE-S | 61 | 13,1 | 21,3 | 42,6 | 8,2 | 8,2 | 0 | 0 |
| Ukrainians | IE-S | 50 | 18,0 | 2,0 | 54,0 | 6,0 | 0 | 0 | 0 |
| Polese | IE-S | 93 | 16,1 | 13,4 | 55,9 | 3,2 | 0 | 0 | 0 |
| Volga-Ural region | |||||||||
| Maris | U-FU | 111 | 8,1 | 2,7 | 47,7 | 31,5 | 9,9 | 0 | 0 |
| Mordvin | U-FU | 83 | 19,3 | 13,3 | 26,5 | 16,9 | 2,4 | 0 | 0 |
| Komis | U-FU | 94 | 5,3 | 16,0 | 33,0 | 22,3 | 12,8 | 0 | 0 |
| Udmurts | U-FU | 87 | 1,1 | 2,3 | 10,3 | 56,3 | 28,7 | 0 | 0 |
| Chuvashes | A-T | 79 | 11,4 | 3,8 | 31,6 | 17,7 | 10,1 | 0 | 1,3 |
| Tatars | A-T | 126 | 4,0 | 8,7 | 34,1 | 18,3 | 4,8 | 0,8 | 1,6 |
| Southern Europe | |||||||||
| Croats | IE-S | 109 | 37,6 | 15,6 | 33,9 | 0 | 0 | 0 | 0 |
| Western Europe | |||||||||
| French | IE-R | 61 | 16,4 | 59,0 | 0 | 0 | 0 | 0 | 0 |
| Central Europe | |||||||||
| Germans | IE-G | 16 | 37,5 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hungarians | U-FU | 113 | 28,3 | 20,4 | 20,4 | 0,9 | 0 | 2,6 | 0,9 |
| Siberia | |||||||||
| Khants | U-FU | 47 | 0 | 19,1 | 4,3 | 38,3 | 38,3 | 0 | 0 |
| Nganasans | U-SA | 38 | 0 | 0 | 0 | 0 | 92,1 | 0 | 5,3 |
| Nenets | U-SA | 148 | 0 | 0 | 0 | 40,5 | 56,8 | 1,4 | 0 |
| Selkups | U-SA | 131 | 0 | 6,1 | 19,1 | 0 | 6,9 | 66,4 | 1,5 |
| Kets | Isolate | 48 | 0 | 0 | 0 | 0 | 0 | 93,7 | 6,2 |
| Dolgans | A-T | 67 | 1,5 | 1,5 | 16,4 | 22,4 | 11,9 | 0 | 37,3 |
| Yakuts | A-T | 155 | 1,3 | 1,9 | 1,9 | 88,4 | 0 | 0 | 3,2 |
| Buryats | A-M | 81 | 0 | 1,2 | 1,2 | 28,4 | 2,5 | 0 | 60,5 |
| Evenks | A-TN | 96 | 5,2 | 0 | 1,0 | 16,7 | 3,1 | 4,2 | 67,7 |
| Evens | A-TN | 31 | 3,2 | 0 | 6,5 | 12,9 | 0 | 0 | 74,2 |
| Altaians | A-T | 98 | 0 | 0 | 46,9 | 1,0 | 2,0 | 17,3 | 22,4 |
| Total | 2967 | ||||||||
В публикации [Balanovsky, 2008] приведены результаты анализа 1228 Y-хромосом русских из 14 регионов России. При учете других данных построены схемы распределения частот гаплогрупп в Европе. Схема частот гаплогруппы N3 показана на рисунке 3. Среди русских число носителей гаплогруппы N3 увеличивается в северо-восточном направлении от 4,5% (Ливни), 6,3% (Репьевка) и 6,6% (кубанские казаки) до 39,5% (Пинега) и 44,4% (Мезень). В этом же направлении уменьшается число носителей гаплогруппы R1a от 62,7% (Ливни), 59,4% (Белгород) до 27,7% (Пинега). Гаплогруппа R1a характерна для восточных славян и поляков. Распределение частот гаплогруппы N3 в Восточной и Центральной Европе соответствует клину (рисунок 3, вставка в верхний сегмент рисунка). Это означает, что в историческое время происходило относительно «плавное» «перемешивание» носителей маркера N3 с носителями европейских маркеров. Этот же массив данных обработан авторами публикации [Fechner, 2008]. Цель обработки – выявление клинов и границ в распределении частот гаплогрупп. Главный клин частот маркера N3 просматривается в Восточной Европе при движении с северо-востока на юго-запад. Еще один клин частот гаплогруппы N3 просматривается при движении с востока на запад: эстонцы – 30,6%, латвийцы – 41,9%, поляки – 3,2%. Среди немцев 1,6% (выборка 1225 человек) носителей этого маркера [Kayser, 2005]. От саамов клин просматривается в южном направлении: саамы – 47,2%, норвежцы – 6,9%, шведы – 2,8%, датчане – 0,5% (таблица 1). У чехов и словаков (45 человек) выявлен 1 носитель маркера N3 [Semino, 2000, The genetic]. На схеме частот гаплогруппы N3 из публикации [Fechner, 2008] (рисунок 3, нижний сегмент) мы условно показали «выклинивания» частот носителей маркера N3 по направлению к территории Венгрии.
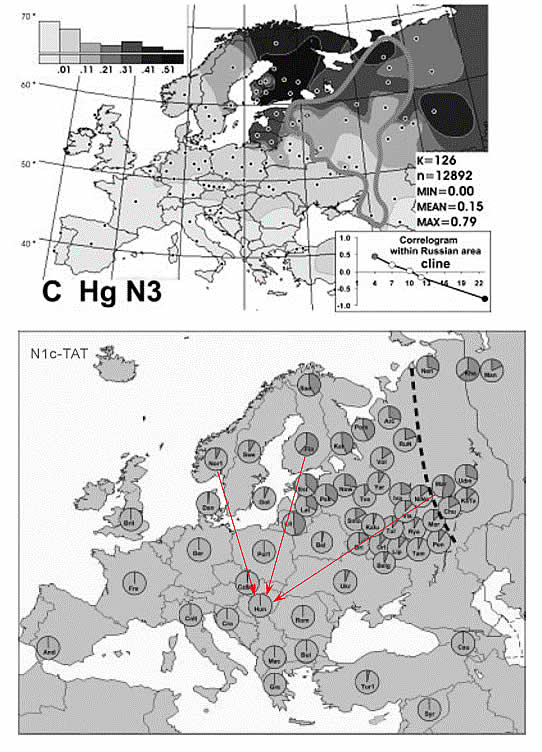
Рисунок 3. Частоты гаплогруппы N3 (N1c-TAT) Y-хромосомы в Европе. Верхний сегмент – [Balanovsky, 2008]. Фигурой неправильной формы обозначена область, охарактеризованная результатами исследований авторов публикации. Нижний сегмент – [Fechner, 2008]. Красными стрелками показаны (А.М. Тюрин) «выклинивания» частот носителей гаплогруппы N3 по направлению к территории Венгрии.
В публикации [Semino, 2000, The genetic] приведены результаты анализа 45 Y-хромосом венгров. Носителей гаплогруппы N3 среди них не выявлено. В другой выборке (количество 113 человек) выявлен один носитель гаплогруппы N3 [Tambets, 2004]. В публикации [Csanyi, 2008] приведены результаты анализа двух выборок мужчин, говорящих на венгерском языке: 100 человек из Венгрии и 97 человек Szeklers (Румыния). Выявлен один (Szeklers) носитель гаплогруппы N3. Еще 2 выборки венгров приведены в публикациях [Battaglia, 2008] (53 человека) и [Volgyi A., 2008] (119 человек). Носителей маркера N3 среди них нет. Автор публикации [Клёсов А., 2009] рассмотрел историю секлеров и ДНК-данные из проекта их изучения. Выборка включает ДНК-данные (гаплогруппы и гаплотипы) по 143 их представителям. У них выявлено 3 носителя гаплогруппы N. Но в тексте статьи мы не нашли пояснений, как она соотносится с гаплогруппой N3. Поэтому данные по этой выборке мы не будем принимать во внимание. Таким образом, в 6 выборках венгров (всего 527человек) выявлено 2 носителя гаплогруппы N3. Это составляет примерно 0,5%. В публикации [Csanyi, 2008] приведены результаты тестирования останков из 4 захоронений на территории Венгрии знатных людей. Тестировалось только наличие гаплогруппы N3. Два теста оказались положительными. То есть в прошлом на Среднедунайской равнине имелись носители гаплогруппы N3, причем они были среди высших слоев общества.
У украинцев носителей маркера N3 6,0%, у поляков их 3,2%, у чехов и словаков – 2,2%. То есть с высокой степенью достоверности можно говорить о том, что маркер N3 попал к венграм от соседей, путем простой эмиссии. Это заключение подтверждается тем, что один из двух венгров, носителей маркера N3 относится к их этнической группе, проживающей в Буковине, то есть в непосредственной близости от Украины. Если не учитывать выборку Szeklers, то среди 258 венгров Венгрии выявлен 1 носитель маркера N3, что составляет примерно 0,4%. Эта ситуация отражена на схеме частот маркера N3 (рисунок 3).
В выборке русских, которую мы уже упоминали (1228 человек), выявлено 8 носителей (0,7%) гаплогрупп C, Q распространенных в Восточной и Северной Азии [Balanovsky, 2008]. Выявлен также один носитель гаплогруппы R2, характерной для юга Азии. В публикации [Roewer, 2008] приведены результаты анализа Y-хромосом русских (545 человек) из 12 областей России (Архангельской, Брянской, Ивановской, Липецкой, Новгородской, Орловской, Пензенской, Рязанской, Смоленской, Тамбовской, Тверской и Вологодской) Выявлено 2 носителя маркера С (по 1 в Рязанской и Смоленской областях) и один носитель маркера R2 (Пензенская область). Всего примерно 0,6%. В таблице 1 приведены частоты гаплогрупп для другой выборки русских (61 человек). Носители маркеров C и Q среди них не выявлены. Не выявлены носители этих маркеров среди украинцев (50 человек) и поляков (93 человека). Среди народов Поволжья маркеры C и Q выявлены только у татар – 1,6%, и чувашей – 1,3% (таблица 1). Всего 3 человека на 580 протестированных представителей народов Поволжья, что составляет 0,5%. То есть в Восточной Европе (за исключением Северного Кавказа, калмыков и башкир, которых мы пока не рассматривали) носителей маркеров, характерных для народов Сибири и Центральной Азии (включая монголов) практически нет. Это ставит крест на одном из самых одиозных элементов ТИ – монгольском завоевании Восточной Европы в 13 веке, с созданием государства Золотая Орда. Но этот вопрос нуждается в отдельном рассмотрении. Это сделано в статье [Тюрин, 2010, Монголы].
В одной из выборок венгров (количество 113 человек) выявлено три носителя гаплогруппы Q и один – C [Tambets, 2004]. В других выборках венгров (45 человек, [Semino, 2000, The genetic], 197 человек [Csanyi, 2008], 53 человека [Battaglia, 2008], 119 человек [Volgyi A., 2008]) носителей гаплогруппы Q и C не выявлено. В выборке, характеризующих секлеров (143 человека) [Клёсов А., 2009], выявлено 4 носителя гаплогруппы Q. Таким образом, среди венгров (количество 670 человек) 1,0% гаплогруппы Q и 0,1% - гаплогруппы С. Здесь несколько настораживает то, что все 7 носителей Q выявлены в 2 выборках. В других выборках их нет. Тем не менее, эти данные следует принять во внимание. Отметим, что 2 носителя гаплогруппы Q выявлены среди молдаван северной Молдавии (выборка 54 человека) [Varzari, 2006]. Вообще, гаплогруппа Q могла попасть к венграм не с востока. Среди евреев ашкенази (выборка 442 человека) выявлено 5,2% носителей гаплогруппы Q [Behar D.M., 2004]. То есть она могла попасть к венграм и молдаванам от евреев. Но это предположение противоречит заключению автора статьи [Клёсов А., 2009], сделанное по гаплотипам секлеров, носителей гаплогруппы Q. Они не соответствуют еврейским.
Данные выборки венгров (45 человек) были обработаны по методу главных компонент [Semino, 2000, The genetic]. В главных компонентах обособлена небольшая группа точек, которые характеризуют Украину, Хорватию, Польшу и Венгрию. Последняя точка находится примерно в центре группы. Анализ частот гаплотипов другой выборки венгров (100 человек из Венгрии и 97 человек Szeklers) показал, что они генетически тесно связаны с популяциями Центральной Европы и Балкан [Csanyi, 2008]. То есть по мужской линии венгры являются типичной центральноевропейской нацией.
3.5. Гаплогруппы мтДНК
По результатам анализа мтДНК101 современных жителей Венгрии и 76 венгров (Sekler) Румынии азиатских маркеров не выявлено [Tomory, 2007]. В другой выборке (98 человек), характеризующей венгров, выявлено по одному носителю восточноазиатских гаплгруп В и М [Semino, 2000, MtDNA]. Сделаны предположения о том, что они не принадлежат предкам венгров, пришедших с Урала. Среди другой группы венгров (55 человек) выявлено 3 носителя гаплогруппы М [Nadasi, 2007]. В публикации [Малярчук, 2008] приведены частоты гаплогруп мтДНК народов, говорящих на финно-угорских языках. Гаплогрупп В и М среди них нет. Для русских приведены данные по четырем выборкам (201, 456, 228 и 134 человека). Три носителя гаплогруппы М (1,5%) выявлено только в первой выборке. Носители гаплогруппы М присутствуют среди чехов (3 человека из 279), поляков (8 человек из 436) и боснийцев (2 человека из 144). Среди словенцев (104 человека) их нет. Носители гаплогруппы В среди перечисленных выборок славянских народов нет. Можно принять версию авторов публикации [Semino, 2000, MtDNA]. Носители гаплогруппы М в двух выборках, характеризующих венгров, имели цыганских предков по материнской линии. Носитель гаплогруппы В одной из выборок приобрел ее не от предков, пришедших из-за Урала. Таким образом, среди венгров не выявлены гаплогруппы мтДНК, характерные для сибирских нардов и монголов. Это относится к современным венграм. По результатам анализа мтДНК 27 останков из захоронений 10-11 веков на территории Венгрии [Tomory, 2007] 2 уверенно отнесены к азиатским, остальные – к западноевропейским. То есть, азиатские гаплогруппы на территории Венгрии были.
Наш вывод об отсутствии среди венгров гаплогрупп мтДНК, характерных для сибирских нардов и монголов, не соответствует выводу авторов публикации [Малярчук, 2008] о наличии этих маркеров у венгров, чехов, поляков, боснийцев и русских. Однако этот вывод основан на «цыганской» гаплогруппе М. На ней же основано «подтверждение» генетическими данными представлений ТИ о существовании в прошлом волн «миграций целого ряда степных народов (гунны, авары, булгары, монголы)» которые «прокатились по Восточной Европе». Похоже, цыгане являются единственным генетическим «подтверждением» «монголоидных» волн с востока. Если же говорить о гаплогруппах, распространенных в Сибири и Центральной Азии, то среди чехов выявлено два их носителя (N9a и А) [Малярчук, 2008]. Среди поляков, боснийцев, словенцев и русских (201 человек) их не выявлено. В последней публикации приведены данные и по русским. Частоты монголоидных гаплогрупп русских центральных и южных регионов (458 человек) – 1,3%, северо-западного региона (228 человек) – 4,0%, поморов (134 человека) – 5,2%. Но это с учетом гаплогруппы M*, которая, как мы поняли, является одной из модификаций «циганской» гаплогруппы М. О «цыганском источнике» гаплогруппы M* говорит тот факт, что ее нет у карелов (83 человека, 4,8% восточноевразийских гаплогрупп), финнов (580 человек, 1,8%), мордвы (102 человек, 2,9%), удмуртов (101 человек, 20,8%) и коми (136 человек, 11,8%). Этой гаплогруппы нет и у поморов. Но она имеется у марийцев (136 человек, 7,4%). В целом, частоты мтДНК восточноевразийских гаплогрупп у русских коррелируются с частотами у них гаплгруппы N3. Это однозначно исключает «монгольский» источник восточноевразийских гаплогрупп мтДНК у русских.
На форумах сайта «Славянская антропология» [http://slavanthro.mybb3.ru/index.php] в развитие темы «Венгры: генетика, лингвистика, история» дана справка по венграм и словакам из недоступных нам баз генетических данных. В выборке по венграм (526 человек) выявлено 6 (1,1%) достоверно восточноевразийских (не цыганских) маркеров. В выборке по словакам (510 человек) их 9 (1,8%). Эти данные мы примем во внимание.
3.6. Обобщение генетических данных по венграм
По общегенетическим характеристикам, гаплогруппам Y-хромосом и мтДНК венгры являются типичной центральноевропейской нацией. Сибирские и центральноазиатские маркеры у них практически отсутствуют. Предки венгров, пришедшие на Среднедунайскую равнину с востока, не являлись социальными общностями, родственными предкам сегодняшних сибирских (включая хантов и манси) и центрально-азиатских (включая монголов) народов.
При таких жестких выводах, согласовать исторические и генетические данные, характеризующие венгров, можно на основе одной из двух посылок. На Среднедунайскую равнину с востока:
- никто не приходил, а венгры как нация, сформировались на основе только автохтонного субстрата;
- пришли социальные общности, генетический портрет которых не выделяется на фоне генетического портрета современных венгров.
Сегодняшний язык венгров, относящийся к финно-угорской группе, а так же относительно достоверные свидетельства о том, что предки их двух этнических групп говорили до 16 века на тюркском, а одной – на иранском языках, практически однозначно исключают вариант «никто не приходил». То есть мы решили первую конкретную задачу: на Среднедунайскую равнину пришли социальные общности, генетический портрет которых имел те же характеристики, что и генетический портрет ее автохтонного населения. Значит, задача дальнейшего поиска предков венгров пришедших с востока формулируется предельно просто. Кто мог придти на Среднедунайскую равнину с востока и при этом имел генетический портрет, близкий к генетическому портрету современных венгров?
4. Лингвистический аспект
Лингвистический аспект решаемой задачи рассмотрен в статье [Тюрин, 2010, Венгры, Лингвистика]. Общий вывод однозначен. Слова, которыми называли и называют венгров и их этнические группы, а также народы и племена, говорящие на языках уральской семьи, прямо и однозначно относятся к лингвистическим полям, которые мы ранее назвали «Лингвистические маркеры Руси-Орды» и «Лингвистические проявления дохристианских культов». Общая реконструкция тоже однозначна. На рубеже 13-14 веков из области восточнее Днепра на Среднедунайскую равнину перешли хорошо организованные ОРДы, включающие мужчин-воинов, их семьи и скот. В иранских, арабских и византийских свидетельствах они фигурируют под именем МАДЖАРы и МАДЬЯРы. В западных источниках – КУМАНы. Эти названия отражают социальную организацию ОРД, основой которой был АЙМАК. В связи с этим ОРДы называли также МОГОЛами и МОНГОЛами. В русских свидетельствах эта социальная общность фигурирует под названием УГРы. Это слово является синонимом слова ОРДА. Возможно, появление УГРов/МАДЬЯР на Сренедунайской равнине явилось результатом Троянской войны 13 века [Фоменко, 2005, Методы]. Возможно, это связано с первым этапом формирования Империи Русь-Орда. В соответствии с [Носовский, Фоменко, 2005, Империя] ее создание в самом начале 13 века началось с района Нижней Волги. Но после создания Империи, МАДЬЯРы Северного Причерноморья и Среднедунайской равнины стали воинами Руси-Орды. Их подразделения получили знамена с изображением волчьей головы. После этого МАДЬЯР стали называть в арабских и персидских свидетельствах и БАШКИРами, ЯСами, ЯЗЫГами, а на самом западе ареала их расселения (западная Чехия) – ПСОГЛАВцами. Часть МАДЬЯР, воинов Руси-Орды после создания Империи была поселена на Среднедунайской равнине в качестве оккупационных войск, которые контролировали Центральную Европу. МАДЬЯРы были адептами фаллистического культа, в котором главный элемент поклонения назывался словом ЯН. Поэтому их называли в свидетельствах и как ХУНы, КУНы, ГУННы и ВЕНгры. Печенеги – это тоже одно из названий воинов Руси-Орды. Этот вопрос рассмотрен в публикации [Тюрин, 2008, Волк и Русь-Орда].
5. Генетические данные по древним жителям Южной Сибири
В публикации [Keyser, 2009] приведены результаты генетического анализа образцов останков из курганных захоронений Южной Сибири (верхнее течение Енисея, Минусинская котловина). 10 из них характеризуют андроновскую археологическую культуру (1800-1400 годы до н.э.), 4 –карасукскую (1400-800 годы до н.э.), 12 – тагарскую (800 лет до н.э. – 100 год н.э.) и 6 – тыштыкскую (100-400 годы н.э.). Андроновская и карасукские культуры относятся к бронзовому веку, две других – к железному. Тагарская культура считается скифской, одновозрастной и однотипной со скифской культурой Северного Причерноморья.
Удалось идентифицировать гаплогруппы 10 Y-хромосом и 26 мтДНК останков людей. Девять Y-хромосом относятся к гаплогруппе R1a1, одна – к С(хС3). Сегодня максимальные частоты гаплогруппы R1a1 (более 50%) отмечены у южных алтайцев, киргизов, русских (центр и юг европейской части России), украинцев, поляков и некоторых групп таджиков. Гаплогруппа С(хС3) характерна для сибирских народов. 20 мтДНК (77%) относятся к западноевразийским гаплотипам (HV, H, T, I, U и K), 6 (23%) – к восточноевразийским (C, Z, G2a, F1b и N9a). Причем, для культур бронзового века доля первых составляет 90%, для железного – 67%. Выполнен и анализ генов, отвечающих за некоторые антропологические характеристики людей. Установлено, что люди, останки которых изучены, были голубоглазые (или зеленоглазые), светлокожие и светловолосые [Bouakaze, 2009].
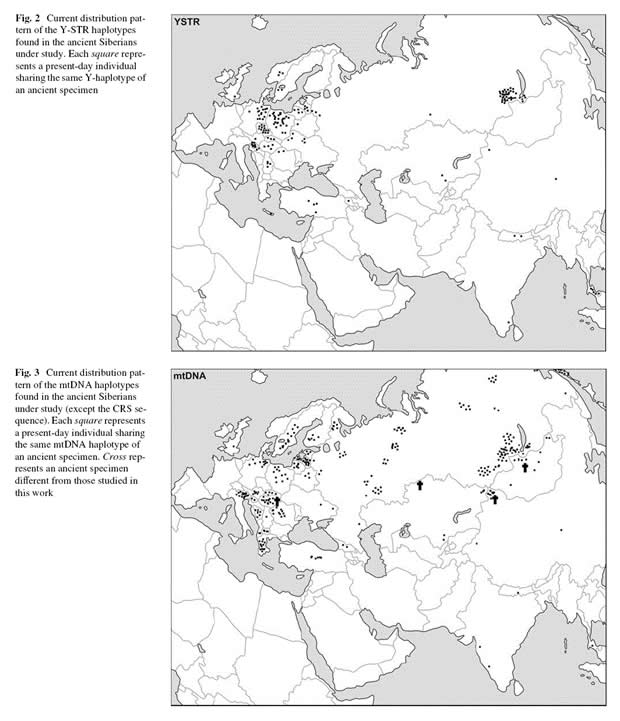
Рисунок 4. Распределение гаплогрупп Y-хромосомы (верхний сегмент) и мтДНК (нижний сегмент) современных популяций, соответствующих гаплогруппам останков из курганных захоронений Южной Сибири [Keyser, 2009].
Авторы публикации [Keyser, 2009] не остановились на тех результатах, которые мы привели выше. Они дали в своей статье таблицы соответствий гаплотипов образцов останков древних людей и гаплотипов из базы генетических данных. Для Y-хромосы аналогов единственного носителя гаплогруппы С не найдено. Для двух гаплотипов гаплогруппы R1a1 в Венгрии найдено 6 аналогов. Для 5 гаплогруп мтДНК (все западноевразийские) найдено 21 аналог гааплотипов, причем, один из них оказался не современным, но из захоронения 11 века на территории Венгрии. Этот образец из той выборки, сведения по которой мы привели выше [Tomory, 2007].
6. Реконструкции
6.1. Идеологическое поле геногенеалогических и геногеографических реконструкций
В соответствии с Новой Хронологией А.Т. Фоменко и Г.В. Носовского (НХ ФиН) [Сайт Новая Хронология], так называемая монгольская империя – это Русь-Орда, возникшая в 14 веке в Восточной Европе и начавшая отсюда экспансию на восток, юг, и запад. Военным инструментом экспансии являлась Орда – армия, сформированная и организованная на основе определенных принципов [Носовский, Фоменко, 2005, Новая хронология Руси]. В своих реконструкциях авторы НХ ФиН четко обозначили несколько моментов, важных для нашей темы. Они дали новое смысловое наполнение применяющегося в исторических хрониках термина СКИФЫ [Носовский, Фоменко, 2005, Империя]. Скифами в свидетельствах называли жителей митрополии Руси-Орды, а за ее пределами воинов Орд. Рассмотрели некоторые вопросы соотношения СКИФОВ НХ ФиН и того, что понимается в Традиционной Археологии под скифской археологической культурой [Носовский, Фоменко, 2005, Империя; Носовский, Фоменко, Античный скиф]. Выявили существование в прошлом в Саяно-Алтайском регионе самой восточной Орды, называвшейся ПЕГОЙ. Предположили, что изгнанные войсками Романовых с Нижнего Поволжья руководители одного из пост-Ордынских формирований набрали в Пегой орде войско и в 40-х годах 17 века завоевали Китай. В ТИ эта акция «замаскирована» под завоевание Китая манжурами [Носовский, Фоменко, 2005, Империя].
6.2. Свидетельства о воинах верховьев Енисея и Приангарья
В [Носовский, Фоменко, 2005, Новая хронология Руси] со ссылкой на работу по истории Сибири приведены следующие данные. «В половине ДВЕНАДЦАТОГО ВЕКА в восточной и центральной Азии жили самостоятельные племена, носившие название КАЗАЧЬИХ ОРД. Наиболее значительная «казачья орда» жила в верховьях реки Енисея и занимала земли на востоке от озера Байкал и на западе до реки Ангара. В китайских хрониках орда эта называлась хакассы, что по исследованию европейских ученых равнозначно слову «казак». По запискам, оставленным современниками, – «хакасы» или «казаки» принадлежали к индо-иранской расе. Они были белокуры и светловолосы; высокие ростом, с зелено-голубыми глазами; храбры, горды и в ушах носили кольца. (Рихтер, немецкий историк 1763-1825 гг., «Иохим», Записки о Монголии).». Специалисты, реконструировавшие по генетическим данным облик людей, чьи тела покоились в курганных захоронениях верховий Енисея, практически дословно повторили это свидетельство «south Siberians were blue (or green)-eyed, fair-skinned and light-haired people» [Keyser, 2009]. То есть потомки скифов-тагарцев благополучно дожили до этнографического этапа изучения народов Сибири и при этом сохранили голубые и зеленые глаза, а так же светлые волосы. Такого, конечно, не могло быть в том случае, если тагарцы исчезли в начале 1 тысячелетние н.э. Не могла сохраниться и народная память о них, если они жили в «половине ДВЕНАДЦАТОГО ВЕКА». Они жили в регионе и в середине 18 века, продолжая хоронить тела умерших сородичей в родовых курганах. Именно эти курганы археологами датированы 1 тысячелетием до н.э.
«Использовали латы и буряты, в донесении из Верховенского острога в 1645г. сообщалось: «А приходят де братские люди войною под острог на конях збройны, в куяках с наручи и в шишаках» (Батуев,1996, с. 74). В 1688 г. иркутский воевода А. Синявин предписывал своим порученцам взять «...братских людей по выбору и развесть в Ыркуцкий сто пятьдесят тотчас с коньми и с ружьем, саадаки, и с куяки, и с панцири» (Михайлов,1993,с.42). В донесениях в Москву особо подчеркивалось высокое качество бурятских доспехов: «...брацкие мужики... бывают на боях в куяках и в наручах, и в шишаках («А мы» – Л.Б.) и со своих худых пищаленок их брацких куяков не пробиваем». Судя по русским письменным источникам «брацкие люди» продолжали применять «панцири и куяки» и во второй половине XVIII в.» [Бобров, 2000]. Братский острог на Ангаре (сегодня город Братск) получил свое название от «братских мужиков». Конечно они не буряты. В публикации [Заилийский Алатау] отмечается, что на южных склонах Заилийского Алатау кочуют «буруты, или дикокаменные киргизы». В 1688 году «дикокаменные киргизы» – «буруцкие люди» (буруты) участвовали в войне на Алтае [Уманский]. Этот вопрос мы подробно рассматривали в публикации [Тюрин, 2008, Волк и Русь-Орда]. Но здесь дадим другую трактовку этого слова. БРАЦКИЕ (БР+ИЦ+аК) люди – это воины. БОРЕЦ – воин. Воинское соединение БОРЦев будет называться БОР+УД. Воины этого соединения могут называться БОР+УДы или БОР+УД+ИШ. Это и есть БУРУТы Заилийского Алатау и БУРТАСЫ Средней Волги. Это и БРИТТы, воины Руси-Орды, завоевавшие в 14 веке Англию. Отсюда БРАТ и BROTHER, слова, первоначально означавшие «сослуживец». То есть в 17 веке сибирские воеводы в Приангарье столкнулись с воинами пост-Ордынского формирования, которое ранее входило в Пегую орду. Важно то, что эти воины были вооружены лучше, чем казаки правительственных войск. Важно и то, что их брали на государственную военную службу. Предками «буруцких людей» были тагарцы. Поэтому их потомки, живущие в Приангарье, дали массовое совпадение генетических маркеров по мужской линии с маркерами останков из курганных захоронений.
6.3. Общая реконструкция
В статье [Тюрин, 2010, Венгры, Лингвистика] мы восстановили несколько дат, характеризующих предков венгров, но не настаиваем на их высокой достоверности. Будем относиться к ним как к условным. Пегая орда поставляла воинские орды для завоевательных походов Руси-Орды. Начало этой практики относится к завершающему этапу Троянской войны, то есть к концу 13 века. Эти орды известны в Европе под названием ПОЛОВЦы, КИПЧАКи, ПЕЧЕНЕГи, КЕЛЬТы, КУМАНы, АВАРы, МАДЬЯРы, МОГОЛы, МОНГОЛы, ТУРКи, …. После завоевания Европы Русью-Ордой в первой половине 14 века, пегие орды были оставлены на территории Центральной Европы в регионе, где сегодня наблюдается максимальная частота генетических маркеров, совпадающих с маркерами останков из курганных захоронений Южной Сибири. На рисунке 5 границы региона, в котором оставлены войска, сформированные в Пегой орде, обозначены пунктирной линией красного цвета. В своей массе люди, пришедшие в Европу из Саяно-Алтайского региона, были носителями западноевразийских гаплогрупп. Поэтому подавляющее число их потомков по гаплогруппам не отличается от потомков автохтонного населения Европы. Потомки пегих орд в Европе были выявлены по результатам сопоставления генетических маркеров останков из захоронений Южной Сибири с данными из генетических баз. Но пегие орды принесли с собой в Европу и восточноевразийские гаплогруппы. Сегодня количество носителей гаплогрупп, характерных для сибирских и центрально-азиатских народов составляет в Восточной Европе (без учета башкир, калмыков и народов Северного Кавказа) менее 1,0%. На территории Центральной Европы рассматриваемые гаплотипы выявлены в женской линии (порядка 1,0% процента), но практически отсутствуют в мужской (кроме венгров).
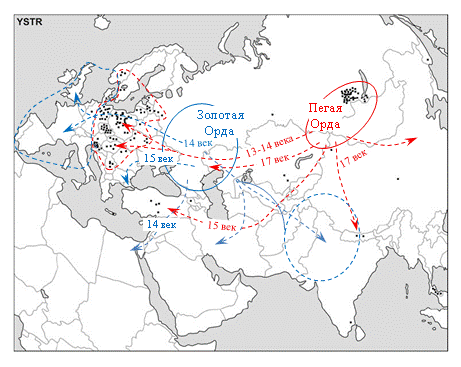
Рисунок 5. Реконструкция активности Пегой орды в период конец 13 – первая половина 17 веков.
Основа схемы соответствует верхнему сегменту рисунка 4. Пояснения даны в тексте статьи.
Пегая орда поставляла орды для завоевательных походов Руси-Орды и в 15 веке. Ранее нами рассмотрены [Тюрин, 2009, Гордион] погребальные сооружения Гордиона, относимые в ТИ к 1 тысячелетию до н.э. Профессиональными археологами показано [Marsadolov, 2000], что погребальные сооружения Гордиона принципиально идентичны скифским курганам. По результатам ре-фальсификации радиоуглеродных дат, характеризующих объекты Гордиона, восстановлена хронология связанных с ним событий. Появление в Анатолии одного из подразделений Орды в качестве оккупационных войск отнесено примерно к 1450 году. Это наше заключение подтверждено результатами анализа генетических данных [Keyser, 2009]. Примерно в районе Гордиона выявлено несколько носителей маркеров Y-хромосомы и мтДНК, идентичных маркерам останков из курганных захоронений Южной Сибири (рисунки 5).
В публикации [Cinnioglu, 2004] приведены результаты изучения Y-хромосомы 523 турок. Образцы отобраны в 9 регионах Турции. Всего выявлено 3,4% (по нашему расчету – 3,1%) носителей гаплогрупп, характерных для Центральной Азии (C, Q и O). Число носителей гаплогруппы R1a1 составляет 6,9%, гаплогруппы N3 – 1,0%. По частоте носителей этих маркеров обособились два региона – центральная Анатолия и ее восток (но без Причерноморья). В первом выявлено 3 из 5 (для всей выборки) носителей гаплогруппы N3 и 3 из 10 – гаплогруппы Q. Число носителей маркера R1a1 составляет здесь 5,6%, что соответствует среднему значению для всей выборки. Восточный регион характеризуется повышенными частотами носителей маркеров R1a1 – 11,0%, и Q – 6,1%. В нем сосредоточено 5 из 10 (50,0%) носителей маркера Q и 9 из 36 (25,0%) носителей маркера R1a1. Отметим, что 3 из 5 носителей маркера С3 живут в Стамбуле. То есть частоты рассмотренных выше маркеров подтверждают нашу гипотезу о появлении в центральной Анатолии переселенцев из Саяно-Алтайского региона. В своей массе они были носителями гаплогруппы R1a1, но среди них имелись и носитали гаплогрупп, характерных для центральноазиатских народов. Область проживания переселенцев охватывала и восточную часть Анатолии, возможно и весь Курдистан.
В 17 веке почти все воины Пегой орды ушли в Китай на службу к манжурам. Поэтому среди современного населения Саяно-Алтайского региона не выявлены маркеры мужской линии, характерные для останков из захоронений Южной Сибири. Но маркеры женской линии в регионе сохранились. Они маркируют, в первом приближении, территорию Пегой орды. Гаплогруппа R1a1 Y-хромосомы, являющаяся доминирующей у изученных останков из курганных захоронений Южной Сибири, сохранилась у современных жителей этого региона. Ее носители среди южных алтайцев составляют 53%, северных – 38%. [Харьков, 2007]. Причем в Горно-Алтайске (выборка 20 человек) носителей этого маркера 50,0%. В поселках Бешпельтир (43 человека) и Кульда (46 человек) их 58,14% и 52,13 %. Среди алтай-кижи (южные алтайцы) (92 человека) носители гаплогруппы R1a1 составляют 41,3%, телеутов (47 человек) – 55,3%, хакасов (53 человека) – 28,3%, шорцев (51 человек) – 58,8%, тоджинцев (36 человек) – 30,6%, сойотов (34 человека) – 23,5%, тувинцев (113 человек) – 17,7% [Derenko M., 2006, Contrasting …; Derenko M., 2006, The diversity …]. У телеутов (Bachat Teleuts) Кемеровской области (35 человек) выявлено 31,43% носителей гаплогруппы R1a1 [Харьков В.Н., 2009; Kharkov V.N., 2009]. Среди киргизов ее носители составляют 63% [Wells, 2001], уйгуров – 29%, тувинцев – 14%, монголов и казахов – 4% [Haplogroup R1a (Y-DNA), http://en.wikipedia.org/]. Но абсолютный рекорд частот гаплогруппы R1a1– (82,5%) у этнической группы монголов – Khoton, живущей на северо-западе Монголии [Katoh, 2005]. Про ее предков «смутно» известно только то, что они откуда-то пришли в регион, говорили на тюркском языке и исповедовали ислам. Это и есть один из наборов маркеров воинов Руси-Орды. Сохранились и западноевразийские маркеры мтДНК. Их частота у хакасов – 18,6%, шорцев – 35,3%, алтайцев - 20,7%, тувинцев – 5,6% [Захаров, 2003].
Основные направления завоеваний Руси-Орды, которые осуществлены в 14-15 веках армиями, сформированными на юге Восточной Европы (зона их формирования условно названа «Золотой ордой»), на рисунке 5 показаны синим цветом. Эта схема завоеваний соответствует самым общим реконструкциям создания Империи, выполненным авторами НХ ФиН. Нами предполагается, что оккупационные войска Руси-Орды, оставленные в Западной Европе, Египте, Иране и на Севере Индостана, были сформированы на юге Восточной Европы. Это предположение вполне проверяемо по генетическим данным. Нужно создать массив данных, характеризующих донских и кубанских казаков, а также народы Северного Кавказа (путем выборки из имеющихся генетических данных) и выполнить поиск их аналогов в Евразии и Северной Африке. То есть нужно технически выполнить то, что сделали авторы публикации [Keyser, 2009].
6.4. Предки венгров
До рубежа 13-14 веков автохтонное население Среднедунайской равнины по своему генетическому портрету не отличалось от населения Центральной Европы и сопредельных областей. В нем был высок процент носителей гаплогруппы R1a1, которая являлась доминирующей на территории Польши и Украины. На рубеже 13-14 веков на равнину пришли первые орды, сформированные в Пегой орде и сопредельных с ней территориях. Возможно, они были сформированы из жителей Северного Казахстана и юга Западной Сибири, говоривших на языке финно-угорской группы. Это было время, когда диффузия гаплогруппы N3 с севера только начала достигать этот регион. Поэтому орды, говорящие на протовенгерском языке принесли с собой этот генетический маркера, характерный для всех народов (кроме венгров), говорящих на языках финно-угорской группы, только в мизерных количествах. Два его носителя выявлены по тестированию останков из захоронений на территории Венгрии [Csanyi, 2008]. Но имеются более веские свидетельства проживания в прошлом предков венгров в Северном Казахстане и юге Западной Сибири. В Тургайском регионе Казахстана живет небольшая популяция казахов (порядка 2000 человек), которую называют МАДЖАРы. Их изучили как возможных «родственников» МАДЬЯР [Biro, 2009]. Детально этот вопрос рассмотрен в публикации [Тюрин, 2009, Маджары]. Маджары не являются «родственниками» мадьяр. Но авторы публикации [Biro, 2009] получили интересный частный результат. Среди маджар (выборка 45 человек) выявлен 1 носитель гаплогруппы N/-M231. Можно почти наверняка утверждать, что это линия N3. По результатам поиска гаплотипа гаплогруппы N/-M231 выявлено 40 его совпадений. Наибольшие частоты совпадений получены для Финляндии и Швеции, а также европейской части России. Кроме того, получено 2 совпадения для Szekely, венгров Румынии, и по 1 для жителей Венгрии, Австрии и Белоруссии. Понятно, почему гаплотип гаплогруппы N/-M231 маджар дал наибольшие частоты совпадений с гаплотипами жителей Финляндии. Среди финнов имеется 63,2% носителей гаплогруппы N3 (таблица 1). Понятно и то, что область, в которой находятся гаплотипы-двойники, в первом приближении соответствует области распространения в Европе гаплогруппы N3. Но число совпадений для венгров – 3, при учете небольшого числа среди них носителей гаплогруппы N3 (0,5%) следует признать аномальным. Из этого следует простой вывод. Носитель гаплотипа N/-M231 маджар и часть носителей гаплотипа N3 среди венгров восходят к общему предку. Его локализация очевидна. Это ханты и манси Западной Сибири. Среди хантов, северных соседей казахов выявлено 38,3% носителей гаплогруппы N3 (таблица 1). То есть, версию попадания этого гаплотипа от предков хантов и манси к предкам венгров и маджарам следует считать наиболее вероятной.
Появление орд с востока было следствием бурных событий на Балканах и в сопредельных областях, названных в НХ ФиН Троянской войной. В 14 веке, после создания Империи, на Среднедунайской равнине были оставлены войска Руси-Орды, сформированные в Пегой орде. Они говорили на тюркском языке. Это предки современных этнических групп венгров – Kiskun и Nagykun. На равнину было переведена и одна из орд, которая была расквартирована некоторое время на территории Ирана (или Курдистана). Там она перешла на иранский язык. Это предки этнической группы Jasz. В свидетельствах о прошлом эти орды и воинские отряды Руси-Орды фигурируют под названиями МАДЬЯРы, МАДЖАРы, МОГОЛы, МОНГОЛы, УГРы, БАШКИРы, ПСОГЛАВцы, ЯСы, ЯЗЫГи, ВЕНГРы, ХУНы, КУНы, ГУННы и ПЕЧЕНЕГи. По своей культуре, религии, образу жизни и внешнему виду члены пегих орд кардинально отличались от народов Центральной Европы. Поэтому их появление в регионе современники восприняли как яркое событие и отразили его в своих свидетельствах.
Мужчины пегих орд были в основном носителями гаплогруппы R1a1. Поэтому их потомки не выделяются среди современных венгров. Среди последних по одним данным 60% (выборка 45 человек) носителей гаплогруппы R1a1 [Semino, 2000, The genetic], по другим (выборка 113 человек) – 20,4 % [Tambets, 2004]. Женщины орд тоже были в основном носителями западноевразийских маркеров. Небольшое число (на уровне 1,0%) восточноевразийских маркеров, принесенных пегими ордами, у современных венгров имеется. Наша реконструкция прямо подтверждается генетическими данными. Это выявленное соответствие нескольких маркеров Y-хромосомы и мтДНК на территории Венгрии и маркеров, найденных в останках курганных захоронений Южной Сибири [Keyser, 2009]. Особенно впечатляет соответствие последним, одного маркера мтДНК останков из захоронения на территории Венгрии [Tomory, 2007].
В 15 веке потомки пегих орд Венгрии приняли участие в балканских войнах и завоевании турками Византии. Скорее всего, слово ТУРКи было одним из их названий. Часть уже венгерских участников этих войн осталась на Балканах и в Анатолии. После обособления из Руси-Орды Аттоманской империи, территория Среднедунайской равнины оказалась в ее составе. После поражения в 1683 году турецкой армии около Вены, начался постепенный переход территории равнины под власть Вены.
Мы рассмотрели 8 этнических групп венгров. Но имеется еще одна их группа – МАГЬЯРАБы, живущая на границе Египта и Судана. Их язык отдаленно напоминает венгерский. Считается, что это потомки венгров, переселенных в этот регион в 1517 году турецким султаном Селимом I. Если это так, то султан просто продолжил практику формирования оккупационных войск из воинов Пегой орды и их потомков. Но, возможно, предки МАГЬЯРАБов были воинами Руси-Орды, которые пришли в Египет в 14 веке. Результаты их генетических исследований могут практически однозначно определить, какая из двух версий является верной. Если предки МАГЬЯРАБов пришли с войсками Руси-Орды, то генетический портрет их потомков не будет содержать маркеров, характерных для западноевропейских народов, скандинавов и южных славян. У современных венгров эти маркеры присутствуют с относительно большими частотами.
7. Заключение
Формальная постановка задачи включала следующий вопрос. Имеются ли следы пришельцев на Среднедунайскую равнину с востока в современном генетическом портрете венгров? Ответ на него получен. Да. Это небольшое число носителей гаплогрупп Y-хромосомы и мтДНК (порядка 1%), характерных для сибирских и центральноазиатских народов, а также гаплотипов западноевразийских гаплогрупп, совпадающих с гаплотипами останков из курганных захоронений Южной Сибири.
Источники информации
Бобров Л.А. Защитное вооружение кочевников Центральной Азии и Южной Сибири в период позднего средневековья. Сибирская заимка. №7. 2000. http://www.zaimka.ru/kochevie/bobrov1.shtml Архив 1998–1999 гг.http://www.zaimka.ru/1998-1999/ Сайт «Сибирская заимка». http://www.zaimka.ru/
[Заилийский Алатау] Заилийский Алатау. http://www.brocgaus-book.ru/text/556.htm Энциклопедия Брокгауза и Ефрона http://www.workmach.ru/
Захаров И.А. Центральноазиатское происхождение предков первых американцев. Первые американцы. 2003. № 11. С. 139-144.
http://macroevolution.narod.ru/zaharov_indians.htm Сайт Проблемы эволюции.
http://macroevolution.narod.ru/index.html
Клёсов А. Секлеры восточной Европы и их гаплогруппы и гаплотипы. Вестник Российской Академии ДНК-генеалогии. Том 2, № 1, 2009 январь. Michael Temosh's Storefront. http://stores.lulu.com/temosh
Малярчук Б.А., Перкова М.А., Деренко М.В. К проблеме происхождения монголоидного компонента митохондриального генофонда славян. Генетика. Том 44, №3, 2008.
[Носовский, Фоменко, 2005, Империя] Носовский Г. В., Фоменко А.Т. Империя.Изд. РИМИС. 2005. //chronologia.org/xpon5/index.html Сайт проекта «Новая Хронология». //chronologia.org
[Носовский, Фоменко, 2005, Новая хронология Руси] Носовский Г В., Фоменко А.Т. Новая хронология Руси. Русь. Англия. Византия. Рим. (В трех томах).Изд. РИМИС. 2005. //chronologia.org/xpon4/index.html Сайт проекта «Новая Хронология». //chronologia.org
[Носовский, Фоменко, Античный скиф] Носовский Г.В., Фоменко А.Т. Античный скиф Иванько Таруль – 1748 год Нашей Эры. http://chronologia.org/Various/various.htm Сайт проекта «Новая Хронология».
//chronologia.org
[Сайт Новая Хронология] Сайт «Новая Хронология». //chronologia.org/
[Тюрин, 2007, Интеграция, Скифы] Тюрин А.М. Интеграция скифской археологической культуры в Новую Хронологию Фоменко и Носовского. /volume6/tur_int.html Электронный сборник статей «Новая Хронология». Выпуск 6. 2007.
/volume6/index.html Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2007, Датирование, Скифы] Тюрин А.М. Датирование скифской археологической культуры по радиоуглеродным данным. /volume6/tur_datskif.html Электронный сборник статей «Новая Хронология». Выпуск 6. 2007. /volume6/index.html Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2007, Биология, Скифы] Тюрин А.М. Биологические маркеры Руси-орды и скифская археологическая культура. /volume6/tur_biomark.html Электронный сборник статей «Новая Хронология». Выпуск 6. 2007. /volume6/index.html Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2007, Лингвистика, Скифы] Тюрин А.М. Казаки, черкасы, татары, орда и скифская археологическая культура. /volume6/tur_kaz.html Электронный сборник статей «Новая Хронология». Выпуск 6. 2007.
/volume6/index.html Сайт: Новая Хронология.
//chronologia.org/ Татарская электронная библиотека. http://kitap.net.ru/tiurin/1.php
[Тюрин, 2008, Интеграция, Хазары] Тюрин А.М. Интеграция информации по хазарам в Новую Хронологию Фоменко и Носовского.
/polemics/turin_int_hazary.html 2008. Полемика. /polemics/index.html Сайт: Новая Хронология.
//chronologia.org/
[Тюрин, 2008, Волк и Русь-Орда] Тюрин А.М. Волк и Русь-Орда. /polemics/turin_volk_rus_orda01.html Полемика. 2008. /polemics/index.html Сайт проекта «Новая Хронология». //chronologia.org
[Тюрин, 2009, Гордион] Тюрин А.М. Археологическая культура Гордиона (Анатолия) и Новая Хронология Фоменко и Носовского. /volume8/turin_gordion.html Электронный сборник статей «Новая Хронология». Выпуск 8. 2009.
/volume8/index.html Сайт: Новая Хронология.
//chronologia.org/
[Тюрин, 2010, Венгры, Лингвистика]. Тюрин А.М. Анализ лингвистических данных, характеризующих венгров. /turin/
[Тюрин, 2009, Маджары] Тюрин А.М. Маджары Казахстана и их предки (геногенеалогический аспект). /turin/
http://rjgg.molgen.org/index.php/RJGGRE/article/view/25 The Russian Journal of Genetic Genealogy (Русская версия). http://rjgg.molgen.org/index.php/RJGGRE
[Тюрин, 2010, Монголы]. Тюрин А.М. Имеются ли генетические следы монгольских завоеваний 13 века? http://supernovum.ru/public/index.php?doc=113 2009. Supernovum.ru. http://supernovum.ru/ /volume10/turin_mongoly.html Электронный сборник статей «Новая Хронология». Выпуск 10. 2010. /volume10/ Сайт: Новая Хронология. //chronologia.org/
Уманский А.П. Сибирские воеводы о событиях в Центральной Азии в конце XVII столетия. Интернет-портал по истории Алтая. http://new.hist.asu.ru/
[Фоменко, 2005, Методы] Фоменко А.Т. Методы. Издательство РИМИС, Москва. 2005. //chronologia.org/xpon2/index.html Сайт проекта «Новая Хронология». //chronologia.org
Харьков В.Н., Степанов В.А., Медведева О.Ф., Спиридонова М.Г., Воевода М.И., Тадинова В.Н., Пузырев В.П. Различия структуры генофондов северных и южных алтайцев по гаплогруппам Y-хромосомы. Генетика, Том 43, № 5, 2007.
http://medgenetics.ru/UserFile/File/Doc/Evolution%20Doc/kharkov%20Alt2007.pdf Сайт Научно-исследовательского института медицинской генетики Сибирского отделения РАМН. http://medgenetics.ru/
Харьков В.Н., Медведева О.Ф., Лузина Ф.А., Колбаско А.В., Гафаров Н.И., Пузырев В.П., Степанов В.А. Сравнительная характеристика генофонда телеутов по данным маркеров Y-хромосомы. Генетика. Том 45, № 8, Август 2009, С. 1132-1142.
http://www.maikonline.com/maik/showArticle.do?auid=VAFXSOWN9K&lang=ru Электронные версии научных журналов. http://www.maikonline.com/
Хуснутдинова Э.К. Этногеномика и генетическая история народов Восточной Европы. Вестник российской науки. Том 73, № 7, 2003.
http://vivovoco.rsl.ru/VV/JOURNAL/VRAN/03_07/ETHNOGENE.HTM
Balanovsky O., Rootsi S., Pshenichnov A., Kivisild T., Churnosov M., Evseeva I., Pocheshkhova E., Boldyreva M., Yankovsky N., Balanovska E. and Villems R. . Two Sources of the Russian Partilineal Heritage in Their Eurasian Context . The American Journal of Human Genetics, Volume 82, Issue 1, 236-250, 2008. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2253976 PubMed Central. http://www.pubmedcentral.nih.gov/
Battaglia V., Fornarino S., Al-Zahery N., Olivieri A., Pala M., Myres N.M., King R.J., Rootsi S., Marjanovic D., Primorac D., Hadziselimovic R., Vidovic S., Drobnic K., Durmishi N., Torroni A., Santachiara-Benerecetti A.S., Underhill P.A., Semino O. Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe. Eur J Hum Genet. 2009 Jun;17(6):820-30. Epub 2008 Dec 24.
Behar D.M., Garrigan D., Kaplan M.E., Mobasher Z., Rosengarten D., Karafet T.M., Quintana-Murci L., Ostrer H., Skorecki K., Hammer M.F. (2004) Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations. Hum Genet 114:354–365
http://www.familytreedna.com/pdf/Behar_contrasting.pdf Family Tree DNA. http://www.familytreedna.com/
Bouakaze C., Keyser C., Crubezy E., Montagnon D., Ludes B. Pigment phenotype and biogeographical ancestry from ancient skeletal remains: inferences from multiplexed autosomal SNP analysis. Int J Legal Med. 2009 Jul;123(4):315-25. http://www.springerlink.com/content/e48j28227r67184u/ SpringerLink. http://www.springerlink.com/home/main.mpx
Biro A.Z., Zalan A., Volgyi A., Pamjav H. A Y-chromosomal comparison of the Madjars (Kazakhstan) and the Magyars (Hungary). Am J Phys Anthropol. 2009 Jan 23;139(3):305-310.
Brandstatter A., Egyed B., Zimmermann B., Duftner N., Padar Z. and Parson W. Migration Rates and Genetic Structure of two Hungarian Ethnic Groups in Transylvania, Romania. Annals of Human Genetics. Volume 71, Issue 6, 2007.
Cinnioglu C., King R., Kivisild T., Kalfoglu E., Atasoy S., Cavalleri G.L., Lillie A.S., Roseman C.C., Lin A.A., Prince K., Oefner P.J., Shen P., Semino O., Cavalli-Sforza L.L., Underhill P.A. Excavating Y-chromosome haplotype strata in Anatolia. Hum Genet. 2004 Jan;114(2):127-48. Epub 2003 Oct 29. http://hpgl.stanford.edu/publications/HG_2004_v114_p127-148.pdf Human Population Genetics Laboratory. http://hpgl.stanford.edu/
Csanyi B., Bogacsi-Szabo E., Tomory Gy., Czibula A., Priskin K., Csosz A., Mende B., Lango P., Csete K., Zsolnai A., Conant E.K.K., Downes C.S.S., Rasko I. Y-Chromosome Analysis of Ancient Hungarian and Two Modern Hungarian-Speaking Populations from the Carpathian Basin. Annals of human genetics (27 March 2008).
Derenko M., Malyarchuk B., Denisova G., Wozniak M., Dambueva I., Dorzhu C., Luzina F., Miscicka-Sliwka D., Zakharov I. Contrasting patterns of Y-chromosome variation in South Siberian populations from Baikal and Altai-Sayan. Human Genetics. Volume 118, Number 5 / Январь 2006 г. http://springerlink.com/content/l4625753377x621t/ SpringerLink http://springerlink.com/
Derenko M.V., Malyarchuk B.A., Wozniak M., Dambuyeva I.K., Dorzhu C.M., Luzina F.A., Lee H.K., Miscicka-Sliwka D., Zakharov I.A. The diversity of Y-chromosome lineages in indigenous population of South Siberia. Doklady Biological Sciences. Volume 411, Number 1 / Декабрь 2006 г. http://springerlink.com/content/700w6227131q52q7/ SpringerLink http://springerlink.com/
Guglielmino C.R., Judit B., Genetic structure in relation to the history of Hungarian ethnic groups. Human Biology, Jun 1996. http://findarticles.com/p/articles/mi_qa3659/is_199606/ai_n8745452/?tag=content;col1 BNET http://findarticles.com/
Guglielmino C.R., De Silvestri A., Beres J. Probable ancestors of Hungarian ethnic groups: an admixture analysis. Ann Hum Genet. Vol. 64, No. Pt 2. 2000. INTERSCIENCE http://www3.interscience.wiley.com/
Fechner A., Quinque D., Rychkov S., Morozowa I., Naumova O., Schneider Y., Willuweit S., Zhukova O., Roewer L., Stoneking M., Nasidze I. Boundaries and clines in the West Eurasian Y-chromosome landscape: Insights from the European part of Russia. American Journal of Physical Anthropology, 2008. 137(1):41-47.
Kayser M., Lao O., Anslinger K., Augustin C., Bargel G., Edelmann J., Elias S., Heinrich M., Henke J., Henke L., Hohoff C., Illing A., Jonkisz A., Kuzniar P., Lebioda A., Lessig R., Lewicki S., Maciejewska A., Monies D.M., Pawlowski R., Poetsch M., Schmid D., Schmidt U., Schneider P.M., Stradmann-Bellinghausen B., Szibor R., Wegener R., Wozniak M., Zoledziewska M., Roewer L., Dobosz T., Ploski R. Significant genetic differentiation between Poland and Germany follows present-day political borders, as revealed by Y-chromosome analysis. Hum Genet. 2005 Sep;117(5):428-43.
Keyser C., Bouakaze C., Crubezy E., Nikolaev V.G., Montagnon D., Reis T. and Ludes B. Ancient DNA provides new insights into the history of south Siberian Kurgan people. Human Genetics. Saturday, May 16, 2009. http://www.springerlink.com/content/4462755368m322k8/ SpringerLink http://www.springerlink.com/home/main.mpx
Katoh T., Munkhbat B., Tounai K., Mano S., Ando H., Oyungerel G., Chae G.-T., Han H., Jia G.-J., Tokunaga K., Munkhtuvshin N., Tamiya G. and Inoko H. Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis. Gene, Volume 346, 14 February 2005, Pages 63-70. http://www.imbice.org.ar/es/lab_06_b/06.pdf Instituto Multidisciplinario de Biologia Celular. http://www.imbice.org.ar/
Kharkov V.N., Medvedeva O.F., Luzina F.A., Kolbasko A.V., Gafarov N.I., Puzyrev V.P., Stepanov V.A. Comparative characteristics of the gene pool of Teleuts inferred from Y-chromosomal marker data. Russian Journal of Genetics. Volume 45, Number 8 / Август 2009 г.
http://www.springerlink.com/content/c310488576h12187/fulltext.pdf SpringerLink http://www.springerlink.com/
Marsadolov L. The Cimmerian Traditions of the Gordion Tumuli (Phrygia):
Found in the Altai Barrows (Bashadar, Pazyryk). Kurgans, Ritual Sites, and Sttlements:Eurasian Bronze and Iron Age. 2000.
http://www.csen.org/BAR%20Book/BAR.%20Part%2001.TofC.html Center for the Study of Eurasian Nomads. http://csen.org/
Nadasi E., Gyurus P., Czako M., Bene J., Kosztolanyi Sz., Fazekas Sz., Domosi P., Melegh B. Comparison of mtDNA haplogroups in Hungarians with four other European populations: A small incidence of descents with Asian origin. http://www.akademiai.com/content/11762351k2682np7/ Acta Biologica Hungarica. Volume 58, Number 2/June 2007.
Pap M. Population genetic research in Hungary. Acta Biologica Szegediensis. Volume 44(1-4):129-133, 2000. http://www.sci.u-szeged.hu/ABS
Roewer L., Willuweit S., Kruger C., Marion N. Rychkov S., Morozowa I., Naumova O., Schneider Yu., Zhukova O., Stoneking M., Nasidze I. Analysis of Y chromosome STR haplotypes in the European part of Russia reveals high diversities but non-significant genetic distances between populations. International Journal of Legal Medicine, 122, №.3, 2008.
[Semino, 2000, MtDNA] Semino O., Passarino G., Quintana-Murci L., Liu A., Beres J., Czeizel A., and Santachiara-Benerecetti A.S. MtDNA and Y chromosome polymorphisms in Hungary: inferences from the palaeolithic, neolithic and Uralic influences on the modern Hungarian gene pool. European Journal of Human Genetics. Vol. 8, №5. 2000.
[Semino, 2000, The genetic] Semino O., Passarino G., Oefner P.J., Lin A.A., Arbuzova S., Beckman L.E., De Benedictis G., Francalacci P., Kouvatsi A., Limborska S., Marcikiae M., Mika A., Mika B., Primorac D., Santachiara-Benerecetti A.S., Cavalli-Sforza L.L., Underhill P.A. The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective. http://hpgl.stanford.edu/publications/Science_2000_v290_p1155.pdf Science (New York, N.Y.) 2000; 290(5494):1155-9. Human Population Genetics Laboratory (HPGL). http://hpgl.stanford.edu/
Tambets K., Rootsi S., Kivisild T., Help H., Serk P., Loogvali E-L., Tolk H-V., Reidla М., Metspalu E., Pliss L., Balanovsky O., Pshenichnov A., Balanovska E., Gubina M., Zhadanov S., Osipova L., Damba L., Voevoda M., Kutuev I., Bermisheva M., Khusnutdinova E., Gusar V., Grechanina E., Parik J., Pennarun E., Richard C., Chaventre A., Moisan J.-P., Barac L., Pericic M., Rudan P., Terzic R., Mikerezi I., Krumina A., Baumanis V., Koziel S., Rickards O., Stefano G., F., Anagnou N., Pappa K.I., Michalodimitrakis E., Ferak V., Furedi S., Komel R., Beckman L., and Villems R. The Western and Eastern Roots of the Saami—the Story of Genetic «Outliers» Told by Mitochondrial DNA and Y Chromosomes. Am J Hum Genet. 2004; 74(4): 661–682. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1181943 PubMed Central (PMC). http://www.pubmedcentral.nih.gov/
Tomory G., Csanyi B., Bogacsi-Szabo E., Kalmar T., Czibula A., Csosz A., Priskin K., Mende B., Lango P., Downes C.S., Rasko I. Comparison of maternal lineage and biogeographic analyses of ancient and modern Hungarian populations. Am J Phys Anthropol. 2007 Nov;134(3) http://www.ncbi.nlm.nih.gov/pubmed/17632797
Varzari A. Population History of the Dniester-Carpathians: Evidence from Alu Insertion and Y-Chromosome Polymorphisms. 2006. Dissertation der Fakultat fur Biologie der Ludwig-Maximilians-Universitat Munchen. http://edoc.ub.uni-muenchen.de/5868/1/Varzari_Alexander.pdf LMU. http://edoc.ub.uni-muenchen.de/
Volgyi A., Zalan A., Beres J., Chang Y.M. and Pamjav H. Haplogroup distribution of Hungarian population and the largest minority group. Forensic Science International: Genetics Supplement Series
Volume 1, Issue 1, August 2008, Pages 383-385.
http://download.journals.elsevierhealth.com/pdfs/journals/1875-1768/PIIS1875176808000231.pdf ELSEVIER. http://download.journals.elsevierhealth.com/
Wells R.S., Yuldasheva N., Ruzibakiev R., Underhill P.A., Evseeva I., Blue-Smith J., Jin L., Su B., Pitchappan R., Shanmugalakshmi S., Balakrishnan K., Read M., Pearson N.M., Zerjal T., Webster M.T., Zholoshvili I., Jamarjashvili E., Gambarov S., Nikbin B., Dostiev A., Aknazarov O., Zalloua P., Tsoy I., Kitaev M., Mirrakhimov M., Chariev A., Bodmer W.F. The Eurasian Heartland: A continental perspective on Y-chromosome diversity. Proc Natl Acad Sci USA. 2001 Aug 28;98(18):10244-9. http://www.pnas.org/content/98/18/10244.full PNAS http://www.pnas.org/
Дополнительная информация
Wiki: Hungarian people. http://wapedia.mobi/en/Madjars
Пока не найденные статьи
Beer Z., Csete K., Varga T. Y-chromosome STR haplotype in Szekely population. Forensic science international. 2004;139(2-3):155-8.
Egyed B., Furedi S., Padar Z. Population genetic study in two Transylvanian populations using forensically informative autosomal and Y-chromosomal STR markers. Forensic Sci Int. 2006 Dec 20;164(2-3):257-65. Epub 2005 Nov 28.
Volgyi A., Zalan A., Szvetnik E., Pamjav H. Hungarian population data for 11 Y-STR and 49 Y-SNP markers. Forensic Science International: Genetics, Volume 3, Issue 2, Pages e27-e28.
Zalan A., Volgyi A., Jung M., Peterman O., Pamjav H. Hungarian population data of four X-linked markers: DXS8378, DXS7132, HPRTB, and DXS7423. Int J Legal Med. 2007 Jan;121(1):74-7. Epub 2005 Oct 20.
