УДК 575.17
Популяционная генетика: расовая и этническая идентификация киммерийцев, скифов, сарматов, саков и черняховцев
А.М. Тюрин

Ключевые слова: скифы, популяционная генетика, расы, этносы, археология, датирование, Новая хронология.
1. Постановка задачи
По результатам исследований древних ДНК, выполненных в 2017-2018 гг. [Damgaard, 2018; Krzewinska, 2018; Unterlander, 2017], получилось, что скифы раннего железного века являлись конфедерацией племен различного происхождения. В июле 2019 г. опубликована статья с новыми данными по скифам [Järve, 2019]. Ее авторы рассмотрели полногеномные генетические маркеры киммерийской, скифской, сарматской, сакской и черняховской археологических культур степной и лесостепной зон Евразии. Геномы обработаны методом главных компонент, выполнены анализ состава предковых компонентов методом ADMIXTURE и моделирование. Интерпретация полученных результатов осуществлена строго на основе частной модели прошлого Человечества, построенной историками и археологами для рассматриваемого региона. Фактические данные популяционной генетики, по мнению авторов, ее в целом подтвердили. Однако, может быть и другой подход к интерпретации этих данных. Он основан только на достоверной информации, которую можно принять за научные факты. К последним относятся данные популяционной генетики и краниологии, а также свидетельства о событиях в прошлом Человечества, начиная с XVII в. На этой основе нами по полногеномным маркерам выполнена расовая и этническая идентификация части древних геномов. Она, естественно, носит условный характер. Во внимание приняты и гаплогруппы мтДНК скифов и представителей синхронных с ними археологических культур, приведенные авторами рассматриваемой статьи.
2. Экспансия калмыков на запад
Калмыки изначально западные монголы ойраты. В свидетельствах фигурируют и как джунгары. Этапы экспансии калмыков на запад в обобщенном виде приведены в нашей статье [Тюрин, 2017, Калмыки]. В начале XVII в. они вышли с территории западной Монголии и начали продвигаться вниз по Иртышу к границе Московского царства. В 1608 г. отряды калмыков появились на Эмбе и «вступили в контакт» с кочевавшими здесь ногаями Большой орды. В 1613 г. калмыки впервые перешли реку Урал. До начала 30-х годов они оттеснили ногаев с левобережья Волги. В 1639 г. калмыки подошли к Самаре и начали кочевать в ее окрестностях. К этому времени их кочевья заняли всю территорию Большой Ногайской орды. В середине XVII в. калмыки совместно с включёнными в их улусы ногайскими родами Большой орды перешли Волгу. В 1661 г. решился вопрос с ногаями западной части Прикаспия. Они приняли ультиматум калмыков. Часть их мурз стала кочевать вблизи Терека, другая вошла в калмыцкие улусы. В период 1655-1664 гг. Московское правительство заключило с калмыками ряд соглашений. Калмыки вошли в русское подданство и периодически давали шерт (брали на себя определенные обязательства). Им были определены степные угодья. По левому берегу Волги от Каспия до Самары, по правому – до Царицына. На западе – до Дона. Сохранили они и свои кочевья в междуречье Волги и Урала, а также на Эмбе и Иргизе.
У калмыков протестированы дербеты (N=69), торгуты (N=58), хошеуты (N=28) и бузавы (N=52) [Balinova, 2019]. Первые три популяции соответствуют родовым объединениям ойратов, последняя – потомкам калмыков-казаков (донских). Частоты гаплогруппы Y-хромосомы С варьируются от 53,6 до 65,4%, средняя 59,4 %. Частоты гаплогруппы О – от 1,9 до 24,0 %, средняя 4,5 %. Гаплогруппа D в этой выборки калмыков не выявлена. Но ее наличие у этой популяции не вызывает сомнения. Ее частоты у монголов составляют первые проценты. Эти три гаплогруппы мы называем «монгольскими». Они являются генетическими маркерами (от термина «генезис») монголов (калмыков) в Восточной Европе и сопредельных районах Азии. Они же являются генетическими маркерами центрально-азиатской монголоидности в регионах к западу от Центральной Азии. Гаплогруппа С является генетическим маркером собственно монголов. Гаплогруппа О по частоте доминирует у китайцев, гаплогруппа D – у корейцев и японцев. Эти две гаплогруппы попали к монголам от соседних популяций.
По результатам авторской интерпретации данных популяционной генетики [Тюрин, 2010; 2017, Литовские татары; 2017, Казахские торе; 2017, Ногаи, генетика] и антропологии [Тюрин, 2018, Ногаи, антропология] при учете достоверных археологических фактов [Матюшко, 2011] и исторических свидетельств [Трепавлов, 2016] сделан категорический вывод: калмыки были первой популяцией, которая принесла в Восточную Европу и сопредельные районы Азии центрально-азиатскую монголоидность и ее индикаторы – гаплогруппы Y-хромосомы C, O и D. Этим самым сформирован новый естественнонаучный метод изучения популяций – датирование по монголоидности. Краниологические серии с признаками монголоидности принадлежат ногаям, метисированным при контактах с калмыками. Их и могильники, по которым они сформированы, в регионах к востоку от Волги следует датировать периодом не ранее второй четверти XVII в., а западнее – не ранее середины XVII в. (время прихода калмыков в обозначенные регионы). По могильникам датируются археологические культуры.
Датированием по монголоидности получены следующие результаты.
1. Верхний хронологический рубеж популяций и могильники, относимых к бронзовому веку, в Среднем Заволжье – середина XVIII в. н. э., популяций на территории Казахстана – начало XVII в. н. э., синташтинской культуры – начало XVII – середина XIX вв. н. э. [Тюрин, 2018, Монголоидность, бронзовый].
2. Верхний хронологический рубеж сарматской культуры Южного Урала и Нижней Волги – XVIII в. н. э. Относимые к ней курганные погребения принадлежат ногаям, а в Южном Зауралье, возможно, казахам. Серии, относимые к позднесарматскому этапу Волго-Уральского региона с признаками монголоидности принадлежат джембуйлукам и джетисанцам Большой ногайской орды [Тюрин, 2018, Монголоидность, сарматы].
3. Хазары-монголоиды датируются периодом не ранее середины XVII в. Идентифицируются они тоже однозначно – это калмыки и ногаи, в предках которых были калмыки [Тюрин, 2017, Монголоидность, хазары].
Выше обозначены индикаторы центрально-азиатской монголоидности по гаплогруппам Y-хромосомы (передаются по мужской линии) – гаплогруппы C, O и D. У гаплогруппы N имеются восточноевразийские линии. Они связаны, главным образом, с популяциями Восточной Сибири. В гаплогруппах мтДНК (передаются по женской линии) выделяются западно- и восточноевразийские линии. Следы экспансии калмыков на запад наглядно проявились в результатах кластеризации полногеномных маркеров (передаются по мужской и женской линиям) выборки популяций Евразии [Yunusbayev, 2015]. Выделено восемь кластеров (Рис. 1). Они уверенно идентифицируются: Европа (К1), Средний Восток (К2), Кавказ (К3), Южная Азия (К4), Сибирь (К5, К6, К7), Восточная Азия (К8). Маркеры кластеров К6 и К8 можно условно назвать «монгольскими». Они резко доминируют у калмыков – 75 %. У них же заметная доля маркеров кластеров К1 и К3. Это результат метисации калмыков при их контактах, главным образом, с ногаями. Маркеры кластеров К6 и К8 доминируют у киргизов, казахов и каракалпаков. Эти популяции сформированы в том числе и на основе калмыков. Заметная доля маркеров кластеров К6 и К8 у популяций Средней Азии – узбеков, таджиков и туркмен. Следы экспансии калмыков у других популяций рассмотрены ниже.
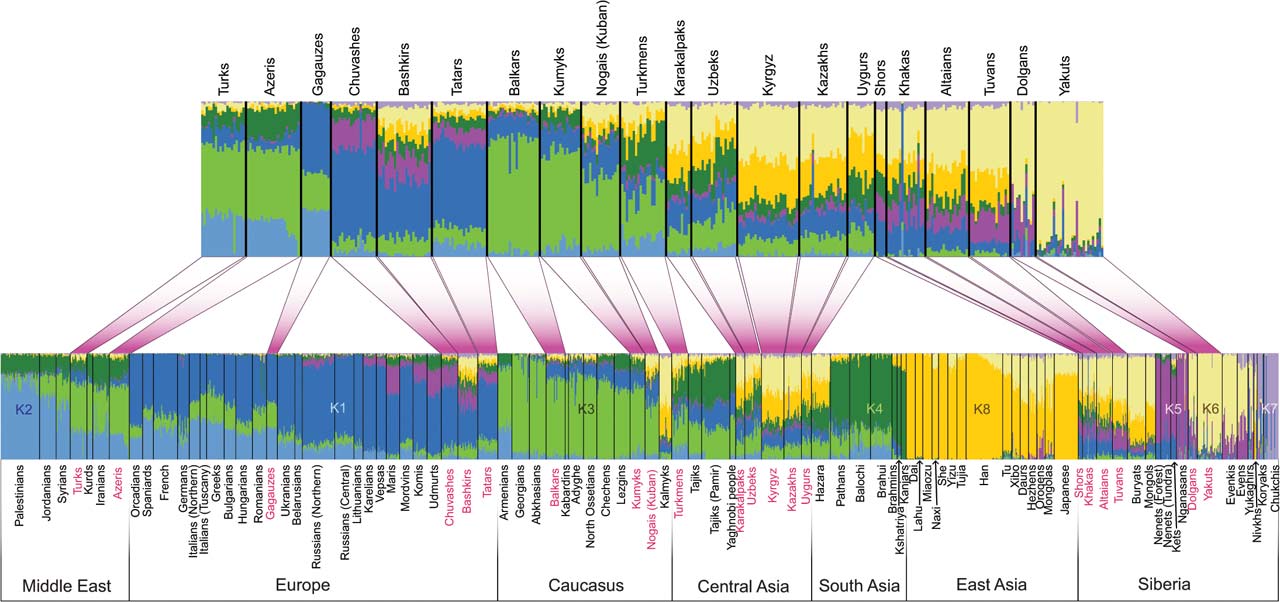
Рис. 1 – Структура полногеномных данных популяций Евразии выборки авторов публикации [Yunusbayev, 2015, Fig. 2]
3. Современные популяции
На рисунке 2 приведена диаграмма главных компонент полногеномных маркеров [Järve, 2019]. Выборка включает современные популяции (приведены медианы точек полногеномных маркеров) и индивидуальные точки, характеризующие останки представителей археологических культур. Современные популяции локализованы на диаграмме на удивление логично. Ее горизонтальная ось соответствует направлению с запада на восток, а вертикальная – с юга на север. В крайней левой части диаграммы снизу-вверх: популяции Ближнего Востока и Закавказья, сардинцы, тосканцы, северные итальянцы, баски Франции, французы, оркнейцы, шведы, поляки литовцы и латвийцы. В правой – китайцы, японцы, нанайцы, орогены и эвены. В самой верхней части диаграммы – саамы.
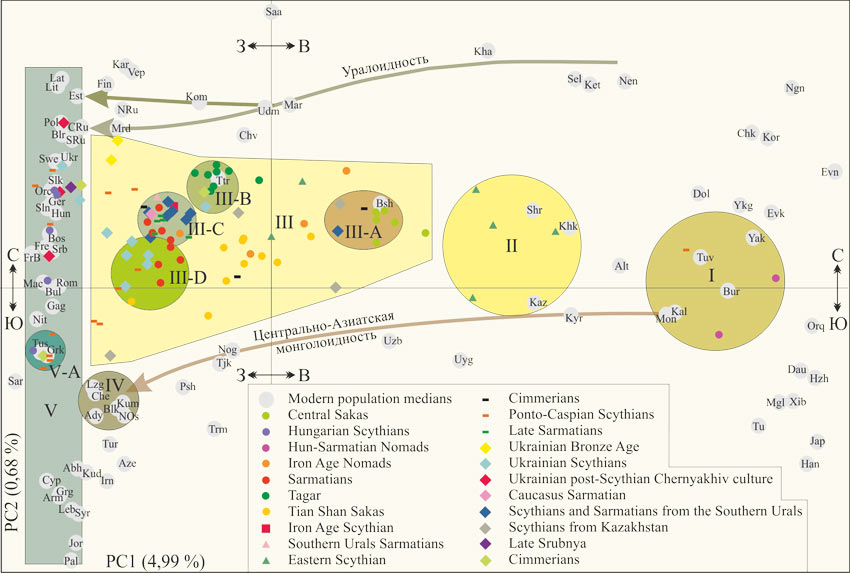
Рис. 2 – Диаграмма главных компонент полногеномных маркеров современных популяций Евразии (показаны медианы) и древних геномов [Järve, 2019]. Интерпретация (кластеры и стрелки потоков генов) А.М. Тюрина
Европа: Bsh,башкиры; Blr, белорусы; Bos, боснийцы; Bul, болгары; Chv, чуваши; Cyp, киприоты; Est, эстонцы; Fin, финны; Fre, французы; FrB, баски Франции; Gag, гагаузы; Ger, немцы; Grk,греки; Hun, венгры; Kar, карелы; Kom, коми; Lat, латвийы; Lit, литовцы; Mac, македонцы; Mar, марийцы; Mrd, мордва; Nit, северные итальянцы; Orc, оркнейцы; Pol, поляки; Rom, румыны; NRu, северные русские; CRu, центральные русские; SRu, южные русские; Saa, саамы; Sar, сардинцы; Srb, сербы; Slk, словаки; Sln, словенцы; Swe, шведы; Ttr, татары; Tus, тосканцы; Udm, удмурты; Ukr, украинцы; Vep, вепсы.
Сибирь: Alt, алтайцы; Bur, буряты; Chk, чукчи; Dol, долгане; Evk, эвенки; Evn, эвены; Ket, кеты; Khk, хакасы; Kha, ханты; Kor, коряки; Mon, монголы; Nen, ненцы; Ngn, нганасаны; Sel, селькупы; Shr, шорцы; Tuv, тувинцы; Yak, якуты; Ykg, юкагиры.
East Asia: Dau, даурцы; Han, китайцы; Hzh, нанайцы (Hezhen); Jap, японцы; Mgl, монголы; Orq, орогены; Tu, Tu; Uyg, уйгуры; Xib, Xibo.
Центральная Азия: Irn, иранцы; Kaz, казахи; Krd, курды; Kyr, киргизы; Psh, пуштуны; Tjk, таджики; Trm, туркмены; Uzb, узбеки.
Ближний восток: Jor, иорданцы; Leb, ливанцы; Pal, палестинцы; Syr, сирийцы; Tur, турки.
Кавказ: Abh – абхазы; Ady – адыгейцы; Arm – армяне; Aze – азербайджанцы; Blk – балкарцы; Che – чеченцы; Grg – грузины; Kal – калмыки; Kum – кумыки; Lzg – лезгины; Nog – ногайцы; Nos – северные осетины.
Центрально-азиатская монголоидность проявилась у популяций в нижней части диаграммы. Кластер I включает популяции, являющиеся эталонными для выделения антропологической монголоидности – монголы, буряты, якуты и тувинцы. В него же попали калмыки. Вблизи границ кластера долгане, юкагиры, эвенки и орогены. Генетические портреты по полногеномным маркерам монголов и калмыков неразличимы. Небольшие различия между ними имеются по частотам гаплогрупп Y-хромосомы.
На диаграмме главных компонент (Рис. 2) по степени убывающей монголоидности популяции ранжированы следующим образом: киргизы, казахи, уйгуры, узбеки, ногайцы, таджики, пуштуны, популяции Северного Кавказа. Монголоидность популяций Северного Кавказа (кластер IV), рассмотрена в наших публикациях [Тюрин, 2019, Караногайцы; 2019, Кубанские ногайцы; 2019, Элементы этногенеза]. «Монгольские» гаплогруппы Y-хромосомы выявлены только у кумыков (N=73) – 1,37 %, и адыгейцев (N=154) – 2,6 % [Кутуев, 2011]. Всего их носителей у пяти популяций (N=154) – 0,7 %. Частоты восточноевразийских гаплогрупп мтДНК у них варьируются от 2,6 (адыгейцы) до 8,3 % (северные осетины). По суммарным частотам гаплогрупп Y-хромосомы и мтДНК популяции ранжированы так: северные осетины (4,1 %), адыгейцы – 2,6 %, балкарцы (2,5 %), кумыки (2,5 %), чеченцы (2,3 %), лезгины (2,2 %). Это в целом соответствует положению характеризующих их точек в поле главных компонент полногеномных маркеров.
Нами установлено [Тюрин, 2019, Кубанские ногайцы], что данные популяционной генетики (четыре группы разных генетических маркеров, характеризующих кубанских ногайцев) из публикаций [Кутуев, 2011; Yunusbayev, 2015] характеризуют, главным образом, ногайцев мансуровцев Малой ногайской орды. По данным популяционной генетики и антропологии они являются типичной популяцией Северного Кавказа. Скорее всего, мансуровцы изначально жили там, где живут сегодня. Помимо северокавказского у них выявлен калмыцкий этнический компонент. Частота «монгольских» гаплогрупп (N=87) – 12,6 %, в том числе: С – 8,1 %, О – 3,4 %, D – 1,1 %. Имеется условно восточноевразийская гаплогруппа N – 4,6 %. У этой популяции (N=131) 20,6 % восточно-евразийских гаплогрупп мтДНК. У мансуровцев (N=16) доля кластеров К6 и К8 составляет 20 %, что согласуется с долей «монгольских» гаплогрупп Y-хромосомы и восточноевразийских мтДНК. В публикации [Järve, 2019] в выборку популяций включены полногеномные маркеры мансуровцев.
Визуально у таджиков доля кластеров К6 и К8 меньше, чем у ногайцев (Рис. 1). Тем не менее, точки, характеризующие эти популяции, практически совпали. Доли этих кластеров у ногайцев и туркмен примерно одинаковые. Это соответствует положению характеризующих их точек на горизонтальной оси диаграммы. Пуштуны получили монголоидность от своих соседей хазарейцев. По частотам у последних доминирую «монгольские» гаплогруппы Y-хромосомы.
После присоединения Крыма к России (1783 г.) часть крымских татар переселилась на территорию Османской империи. Часть ногаев Большой орды была переселена из Прикубанья в Крым [Тюрин, 2017, Калмыки]. Это именно те ногаи, которые пришли на территории западнее Волги в составе калмыцких улусов. В 1857-1861 гг. большая часть ногайцев Северного Кавказа и Крыма эмигрировала в Османскую империю. С Северного Кавказа ушло 30,6 тысяч [Волкова, 1974]. Представление о масштабах переселения из Крыма дает архивный документ, приведенный автором публикации [Алиев, 2017]. Крымские татары Добруджи (часть из них были ногаями) обратились к императору Александру III с прошением о возвращении в Российскую империю. Указана их численность – 70 тысяч. Прошение удовлетворено не было, «многие татары Румынии и Болгарии стали эмигрировать в Турцию» (с. 31). Скорее всего, в Османскую империю существовал и поток калмыков (ясырь).
Генетические следы миграции ногаев («монгольские» гаплогруппы Y-хромосомы и субклад R1b-Z2106) уверенно выделяются у болгар и турок [Тюрин, 2019, Ногаи]. Уверенно они выделяются и по диаграмме полногеномных маркеров (Рис. 2). Заметная монголоидность турок примерно соответствует монголоидности популяций Северного Кавказа (кроме ногайцев). Минимально монголоидность проявились и у других популяций, локализованных на территории Османской империи – курдов и сирийцев. Про источник заметной монголоидности азербайджанцев и иранцев ничего конкретного сказать не можем. Этот вопрос требует специального рассмотрения.
Уралоидность проявилась у популяций, попавших в верхнюю часть диаграммы. Она убывает от кетов, ненцев, селькупов и хантов до центральных русских и эстонцев. Проявилась у популяций Поволжья и финнов, карелов, вепсов, а также северных русских. Это соответствует другим типам генетических маркеров и данным антропологии.
В кластер II попали популяции южносибирской расы – киргизы, казахи, алтайцы, хакасы и шорцы. Это метисы европеоидов и монголоидов. У киргизов и казахов уралоидные компоненты практически отсутствуют. У других популяций они имеются. Границы кластера на диаграмме главных компонент (Рис. 2) обозначены нами с учетом данных по древним популяциям. Кластер V включает европейские и ближневосточные популяции без примеси монголоидности и уралоидности. Эстонцы, центральные русские, абхазы и сирийцы попали на его границу.
Таким образом, на диаграмме главных компонент четко обозначились два потока генов. Первый – монголоидность из Центральной Азии до территорий Турции и Сирии, второй – уралоидность из Восточной Сибири до территории Эстонии. Но две популяции – башкиры и поволжские татары, не вписываются в эту схему. Монголоидные компоненты у башкир мы детально рассмотрели [Тюрин, 2019, Башкиры-1, 2019, Башкиры-2]. В среднем у них 3,0 % «монгольских» гаплогрупп Y-хромосомы С и О (N=935), 39,2 % восточноевразийских гаплогрупп мтДНК (N=666), 20,0% «монгольских» полногеномных маркеров (N=16). Имеются восточносибирские линии гаплогруппы N [Балановская, 2017] (их частоты для популяции в целом оценить мы не можем) и примерно 5,0 % полногеномных маркеров кластера «Сибирь» (Рис. 1). Такой генетический портрет башкир в первом приближении соответствует положению на диаграмме, характеризующей их точки. У них имеется компоненты монголоидности и уралоидности. По татарам полного массива генетических данных у нас не имеется. Скорее всего, у них будет такая же ситуация, как у башкир.
4. Древние геномы
4.1. Европеоиды с минимальными компонентами монголоидности и уралоидности
По основному массиву древних геномов выделен кластер III. В него попали всего две современные популяции – башкиры и поволжские татары. Расовая идентификация кластера однозначна. Это европеоиды с минимальными компонентами монголоидности и уралоидности. Внутри кластера выделены субкластеры. В субкластер III-C включены все скифы и сарматы Южного Урала (N=8), кроме одного, сарматы Южного Урала (N=2), поздние сарматы (N=5), скиф железного века, сармат Кавказа и номад железного века, а также пять сарматов (N=9). Другие четыре сармата попали в субкластер III-D. Он объединяет пять скифов Украины (N=9), два скифа Понто-Каспийского региона (N=14) и одного тянь-шаньского сака. В субластер III-B попали представители тагарской культуры (Минусинская котловина, VIII-III вв. до н. э.) и современные поволжские татары. Скорее всего, это не случайное совпадение. «Тагарское население в целом (включая «грунтовых тесинцев») имеет явное морфологическое своеобразие по сравнению с большинством серий раннего железного века Евразии, демонстрируя в данном масштабе наиболее европеоидный комплекс краниометрических признаков» [Учанева, 2017, с. 84]. Можно предположить, что европеоидный субстрат татар и тагарцев идентичен, а монголоидные и уралоидные компоненты последние получили от своих соседей. Но, конечно, этот вопрос нуждается в специальном рассмотрении. В субкластер III-B также попали один киммериец и один украинский скиф.
В положении трех рассмотренных субкластеров проявилась обозначенная нами закономерность. Уралоидность популяций возрастает по вертикальной оси диаграммы. Минимальная она у популяций на территории степной Украины, максимальная у поволжских татар (Средняя Волга). Популяции Южного Урала занимают промежуточное положение. Эту закономерность подтверждают и современные популяции Северного Кавказа (кластер IV). Проявление уралоидности у них заметно ниже, чем у представителей субкластера III-D.
У скифов Северного Кавказа и Северного Причерноморья (N=24) 20,8 % восточноевразийских гаплогрупп мтДНК (A, C, D, F) [Järve, 2019]. Это однозначно указывает на то, что среди их предков были женщины из Центральной Азии или Восточной Сибири. Среди скифов Южной Сибири (N=24) носители восточноевразийских гаплогрупп (A, C, D, F, G, M, Y, Z) составляют 53, 6 %.
Калмыки были первой популяцией, которая принесла в Восточную Европу и сопредельные районы Азии монголоидность и ее индикаторы – «монгольские» гаплогруппы Y-хромосомы. В соответствии с этим, носители древних геномов, попавшие в кластер III, являются предками башкир и ногаями Большой орды, метисированными при контактах с калмыками. Но показать это на рассматриваемых фактических данных мы не можем. Остановимся на том, что они не противоречат обозначенной концепции.
Ранее мы рассмотрели краниологические серии сарматов [Тюрин, 2018, Монголоидность, сарматы]. Основная часть серий Южного Урала и Нижней Волги принадлежит европеоидам без признаков монголоидности. Они датируются периодом не позднее первой половины XVII в. (до прихода калмыков в обозначенный регион). Вместе с тем имеются серии с признаками монголоидности (могильники Калиновский, Быковский, Политотдельский, Терновский, Кардаилово, Черная 2, Переволочанский, Каменный Амбар и Соленый Дол, погребение близ деревни Черненьки). Они датируются периодом не ранее второй половины XVII в. н. э. (начальный этап метисации ногаев при контактах с калмыками). То есть, по данным краниологии основная часть сарматов не имеет признаков монголоидности. Но в соответствии с нашей интерпретацией результатов обработки полногеномных маркеров все сарматы имели монголоидные компоненты. К этому добавим новые данные популяционной генетики [Järve, 2019]. На гаплогруппы мтДНК тестированы останки ранних сарматов из могильника Покровка (N=11). Носители восточноевразийских гаплогрупп (F, N, M) составляют 27,3 %. Они получены от калмыков. Скорее всего, антропологи «не видят» отражение в характеристиках черепов небольшого содержания монголоидных компонентов. Но один сармат попал в субкластер III-A. Монголоидные компоненты у него на уровне современных башкир. Специалисты такую монголоидность выделяют практически однозначно.
4.2. Киммерийцы
Киммерийская археологическая культура предшествовала скифской. В выборке авторов публикации [Järve, 2019] две группы киммерийцев (одна из публикации [Krzewinska, 2018]), локализованных на территории Украины. Мы не смогли понять, как обозначены характеризующие их точки на диаграмме главных компонент. В одной из групп (N=3): грек и поволжский татарин, третий близок к словакам. В другой (N=3): башкир и тянь-шаньский сак, третий близок к скифам и сарматам Южного Урала. Вывод здесь однозначный: киммерийская археологическая культура (XI-VII вв. до н. э.) не связана с генетически обособленным сообществом (популяцией). Само ее выделение базируется на древнегреческих письменных свидетельствах, в которых фигурируют киммерийцы, и авторитете В.А. Городцова (1860-1945 гг.) [Граков, 1977]. Попросту говоря, данные популяционной генетики закрыли вопрос о киммерийской культуре. Ее не существует.
4.3. Скифы
Группа скифов Южной Сибири (Eastern Scythian) (N=6) разделилась на две части. Четыре ее представителя попали в кластер II. Три из них ближайшие предки шорцев, хакасов и алтайцев, один – казахов. Другая часть группы попала в кластер III между субкластерами III-A и III-B. Можно приять, что это популяция с разной степенью монголоидности ее членов. Четыре ее представителя относятся к южносибирской расе, два – европеоиды с долей монголоидных и уралоидных компонентов заметно меньшей, чем у современных башкир.
Скифы Казахстана (N=4) не являются популяцией. Характеризующие их точки «разбросаны» по кластеру III. Ногаи Большой Орды пришли в регионы восточнее Волги с Северного Кавказа [Тюрин, 2019, Элементы этногенеза]. Поэтому нет ничего удивительного в том, что один скиф близок к популяциям Северного Кавказа (кластер IV).
Пять скифов Украины, локализованы в субкластере III-D, один – рядом с ним, один – украинец, один – вблизи границы кластера V, один попал в субкластер III-B. Можно принять, что эта группа является европеоидной популяцией, в которой большинство членов имеют монголоидные компоненты. Но скифы Понто-Каспийского региона (N=14) однозначно популяцией не являются. Одного представителя группы мы идентифицировали ниже – калмык. Точки, характеризующие других скифов, «разбросаны» на диаграмме по вертикальной оси. Шесть их представителей попали в кластер III. Это европеоиды с минимальной долей монголоидности и уралоидности. Шесть скифов попали в кластер V. Это европеоиды без монголоидных и уралоидных компонентов. Четыре скифа локализованы в субкластере V-A. Это однозначно греки. Три оставшихся – славяне. Но один может быть и венгром. Отметим, что по данным популяционной генетики, в том числе и отраженным на диаграмме (Рис. 2) венгры являются славянами без уралоидных компонентов. Близки к словакам, словецам и боснийцам. То есть, предки венгров не могли прийти в Паннонию с Южного Урала.
Скифы Венгрии (N=5) европеоиды без монголоидных и уралоидных компонентов. Один скиф – грек, один – македонец, румын или болгарин, один – босниец, два – словаки или немцы. Вполне возможно, три последних скифа попадают в «облако» венгров. Скифы Венгрии не являются популяцией.
4.4. Гунно-сарматы
Судя по справочникам и учебникам археологи едины в основных вопросах, связанных с гуннами.
1. Хунну или гунны, – древний центрально-азиатский народ, сформировавшийся в степях Центральной и Северной Монголии, Северо-Восточного Китая, Забайкалья и Южной Бурятии.
2. Гунны яркие участники Великого переселения народов (II в. до н. э. – IV в. н. э.), преобразившие этнополитический ландшафт Евразии от монгольских степей до Придунайских равнин.
3. Внедрившись в среду сарматов Северного Казахстана и Южного Зауралья, гунны создали гунно-сарматское сообщество.
4. Гунно-сарматы были «чистыми» кочевниками (номадами) – повозки, каркасные юрты, минимум посуды и бытовых предметов.
В выборку из публикации [Järve, 2019] включены полногеномных маркеры гунно-сарматских кочевников (N=2) из Южного Зауралья и Северного Казахстана [Damgaard, 2018]. Отметим, что выделение в этих регионах их погребений II-IV в. н. э. носит дискуссионный характер [Мошкова, 2007]. Оба представителя гунно-сарматов попали в кластер I. Причем, монголоидность у них проявилась ярче, чем у монголов и калмыков. Формально, это блестящее подтверждение модели археологов. Кочевники с территории Монголии и сопредельных регионов, пришедшие в Южное Зауралье и Северный Казахстан, просто обязаны быть яркими монголоидами.
По нашей трактовке оба гунно-сармата однозначно идентифицируются как калмыки. Их погребения датируются первой половиной XVII в. По справке П.С. Палласа (по результатам его наблюдений в 70-х гг. XVIII в.) калмыки Северного Прикаспия сжигали тела умерших сородичей, бросали их в степи, в лесу, опускали в воду, просто зарывали в землю или закрывали камнями [Харузин, 1890]. То есть, в регионах проживания в XVII-XIX вв. собственно калмыков их курганных погребений быть не должно. Но с другой стороны есть прямые сообщения о раскопках русскими бугровщиками калмыцких погребений в зоне, примыкающей к укрепленным линиям Южного Зауралья и Западной Сибири. «Штраленберг сообщает, что русские отказывали калмыцким послам в разрешении посетить могилы их предков, опасаясь, что массовое разорение могил вызовет озлобление и вражду со стороны калмыков» [Миллер, 1954]. Скорее всего, специфическая похоронная традиция существовала у некоторых калмыцких родов или социальных групп. В Кластер I попал и один понто-каспийский скиф. Он тоже идентифицируется нами, как калмык. Яркая монголоидность калмыков XVII в. объясняется тем, что они еще небыли метисированы при контактах с ногаями. И, конечно, калмыки являлись «чистыми» кочевниками.
4.5. Центральные саки
Центральным сакам (Central Saks) соответствует тасмолинская археологическая культура Центрального Казахстана. Ее два этапа датируются VII-VI вв. и V-III вв. до н. э. «Новые археологические материалы показывают, что Центральный Казахстан находился в тесных этнокультурных связях с районами Восточного Казахстана, всего Саяно-Алтая, Жетысу, Восточного Приаралья, Южного Урала» [Бейсенов, 2015, с. 12]. На это же указывают данные популяционной генетики. Центральные саки (N=6) неотличимы от башкир, близки к части скифов Южного Урала и Казахстана. Близки к сакам Тянь-Шаня, древним и современным популяциям Алтая. По данным краниологии на диаграмме главных компонент точки «саки Тянь-Шаня», «саки Казахстана (сборная)» и «саки Приаралья (ранние)» находятся экстремально близко [Ходжайов, 2015, рис. 1].
Выделенный нами кластер III- A имеет высокую «плотность». Он включает башкир и центральных саков, по одному скифу Казахстана и Южного Урала, а также киммерийца. Формальный вывод однозначен: в субкластер III-A попали современные башкиры и башкиры XVII-XVIII вв., проживавшие в Центральном Казахстане. Но, возможно, башкиры постоянно проживали на Южном Урале. В Центральном Казахстане были их летние пастбища или зимние охотничьи угодья. Материальное наследие башкир отнесено к тасмолинской археологической культуре.
4.6. Ченяховцы
Готы – участники Великого переселения народов. Они вышли из Южной Швеции (I в. н. э.), оставили материальное наследие на Нижней Висле (вельбарская археологическая культура) и в северо-западном Причерноморье (черняховская культура), сокрушили Римскую империю и создали на ее бывшей территории свои королевства (IV-V вв.). Это основные элементы феномена Традиционной истории «Готицизм».
Образцы (N=3), характеризующие представителей постскифской черняховской культуры (II-IV вв. н. э.), отобраны из погребений на территории Украины на границе степной и лесостепной зон [Järve, 2019]. Ну и кем этнически являются ее представители? Среди них поляк, словак и серб. Шведов не имеется. Это подтверждает наши выводы, сделанные по результатам кластеризации выборки краниологических серий «Восточная Европа, III в. до н. э. – IV н. э.» [Тюрин, 2018, Готы]:
- черняховцы однозначно славяне;
- «Готицизм» – это целенаправленно сконструированная фальшивка.
Здесь интересен один момент. У сербов по частотам доминирует гаплогруппа Y-хромосомы I2 (34 %) [Distribution]. В распределении частот гаплогруппы имеется две зоны повышенных их значений. Одна на территориях Боснии, Хорватии и запада Сербии, другая – на территории Молдовы и сопредельных районов Украины и Румынии. Вторая зона соответствует центральной части территории черняховской культуры. То есть, идентификация одного черняховца как серба подтверждается географией гаплогруппы I2.
Наш вывод определен. Данные краниологии и популяционной генетики являются киллер аргументами против «Готицизма». Но вывод авторов публикации [Järve, 2019, p. 1] диаметрально противоположен нашему: «Our results agree with the Gothic source of the Chernyakhiv culture».
5. Скифская археологическая культура
Приведем точную цитату, характеризующую результаты изучения скифов инструментарием популяционной генетики, выполненного в 2017-2018 гг. «The Early Iron Age nomadic Scythians have been described as a confederation of tribes of different origins, based on ancient DNA evidence» [Järve, 2019, p. 1]. Откуда авторы узнали про конфедерацию племен? Данные популяционной генетики, включая приведенные в статье, на которую ссылка дана выше, свидетельствуют о том, что отдельные группы скифов являются популяциями, другие группы – это объединения генетически разных людей. Никаких сведений о «конфедерации» у генетиков не имеется. Эта нечеткость формулировок обусловлена аберрацией. Генетики действительно считают, что тестировали образцы, характеризующие скифов. Реально они работали с образцами, отобранными из погребений, отнесенных к скифской археологической культуре.
Известный археолог, доктор исторических наук, профессор А.О. Добролюбский «выдал» главную тайну археологии: принцип формирования дубликатных археологических культур путем «расщепления» реальной археологической культуры на части и передатировки одной из них [2000]. Таким образом создаются «ущербные» дубликаты. По результатам профессионально выполненного анализа показано, что, сармато-аланская археологическая культура (I в. до н. э. – IV в. н.э.) северо-западного Причерноморья является сдвинутым в прошлое на 854 года «ущербным» дубликатом печенежско-половецкой культуры (X-XIII вв.).
Ранее мы детально рассмотрели скифскую археологическую культуру (Серия статей «Скифская археологическая культура в Новой Хронологии Фоменко и Носовского», Электронный сборник статей «Новая Хронология», 2007, Вып. 6). Основные результаты изложены в [Тюрин, 2007]. По калибровке радиоуглеродных дат, характеризующих скифскую культуру, по независимой калибровочной кривой [Тюрин, 2005] ее нижний хронологический рубеж 1415-1455 гг. н. э. Это для всего ареала распространения культуры. Верхний хронологический рубеж скифской культуры оценен по комплексу данных. По регионам он разный – 1760 (1850) (Южная Сибирь), 1705 (Алтай), 1800 (Северный Казахстан), 1675 (северное Причерноморье) гг. н. э.
По сегодняшнему нашему пониманию скифская археологическая культура является «ущербным» дубликатом, сформированным по дружинным погребениям времен Империи (XIV-XVI вв.) реконструкций А.Т. Фоменко и Г.В. Носовского [Носовский, Фоменко, 2012], а также погребениям кочевников, продолжавших эту традицию до XIX в. Соответствующую справку привел И.А. Кастанье. «Случалось, даже, что с покойником хоронили его лучшую лошадь. Этот обычай был очень распространен еще в конце XVIII века и в начале XIX по ту сторону Урала» [1911, с. 79]. «По ту сторону Урала» – это Южное Зауралье. Справка относится к казахам. Но для нас важны два момента: в этом регионе проживали и башкиры, а ногаи Большой орды – прямые предки казахов. Курганные погребения с конями – это один из идентификационных признаков дружинных погребений в Восточной Европе и Скандинавии.
Технология формирования «ущербного» дубликата «скифская археологическая культура» рассмотрена нами в публикации [Тюрин, 2007]. К всему многообразию погребений в степной и лесостепной зонах Евразии применен фильтр «скифская триада». Это наличие в них определенных типов конского снаряжения, оружия и предметов, выполненных в зверином стиле. Из погребений, выделенных таким образом, исключены те, в которых имелись явно «не скифские» артефакты. Например, изделия из железа, стремена или венецианское стекло. Оставшиеся погребения отнесены к скифской культуре. В нее попали, главным образом, часть дружинных погребений. Остальные отнесены к скандинавам и славянам IX-X вв., а также золотоордынским кочевникам XIII-XIV вв. Это тоже «ущербные» дубликаты дружинных погребений Империи и постимперских кочевников.
6. Общие выводы
1. На диаграмме главных компонент по медианам полногеномных маркеров современных популяций четко обозначились два потока генов. Первый – монголоидность из Центральной Азии до территорий Турции и Сирии, второй – уралоидность из Восточной Сибири до территории Эстонии.
2. По результатам кластеризации полногеномных маркеров современных популяций и представителей археологических культур выполнена расовая и этническая идентификация последних. Предшествующие скифам киммерийцы обособленной популяцией не являются. Большинство восточных скифов (Южная Сибирь) относится к южносибирской расе. Они ближайшие предки популяций Алтая и казахов. Скифы Северного Причерноморья европеоиды, а также европеоиды с небольшой долей уралоидности и монголоидности. Скифы Казахстана и скифы Понто-Каспийского региона популяциями не являются. Часть последних – греки. Синхронные скифам гунно-сарматы – калмыки, центральные саки – башкиры. Представители постскифской черняховской культуры – славяне.
3. Основная масса представителей археологических культур – киммерийцы, скифы, сарматы и саки, имеют монголоидные и уралоидные компоненты. Это, в основном, ногаи, метисированные при контактах с калмыками. Датируются не ранее XVII в.
4. Идентификация черняховцев как «славяне» является киллер-аргументом против феномена Традиционной истории «Готицизм».
5. Скифская археологическая культура является «ущербным» дубликатом, сформированным по дружинным погребениям времен Империи (XIV-XVI вв.) реконструкций А.Т. Фоменко и Г.В. Носовского, а также погребениям кочевников, продолжавших эту традицию до XIX в. Датируется XV-XIX вв. н. э.
Литература
Алиев Р.Д. Документы Российского государственного исторического архива как источник по истории миграций крымских татар в XIX – начале XX в. // Вестник архивиста, 2017, № 3, с. 22–34.
Балановская Е.В., Юсупов Ю.М., Схаляхо Р.А., Степанов Г.Д., Асылгужин Р.Р., Жабагин М.К., Балаганская О.А, Султанова Г.Д., Борисова Е.Б., Дараган Д.М., Балановский О.П. Генетические портреты семи кланов северо-западных башкир: вклад финно-угорского компонента в генофонд башкир // Вестник Московского университета. Серия XXIII. Антропология, 2017, № 3, с. 94-103.
Бейсенов А.З. Поселения и могильники сакской эпохи Центрального Казахстана // Сакская культура Сарыарки в контексте изучения этносоциокультурных процессов Cтепной Евразии, 2015, с. 11-38.
Волкова Н.Г. Этнический состав населения Северного Кавказа в XVIII – начале XХ века. М., «Наука», 1974, с. 276.
Граков Б.Н. Ранний железный век. М.: Изд-во МГУ, 1977, 235 с.
Добролюбский А.О. Великое перерождение народов. В книге: Г.В. Носовский, А.Т. Фоменко Реконструкция всеобщей истории. Исследования 1999-2000 годов. Москва, изд-во «Деловой Экспресс», 2000, с. 547-562.
Кастанье И.А. Надгробные сооружения киргизских степей. Оренбург: Тип. Оренбургской духовной консистории, 1911, 120 с.
Кутуев, И.А., Хуснутдинова Э.К. Генетическая структура и молекулярная филогеография народов Евразии. Уфа: Гилем, 2011, 239 c.
Матюшко И.В. Особенности погребального обряда кочевников степного Приуралья XIII-XIV вв. // Известия Самарского научного центраРоссийской академии наук, 2011, Т. 13, № 3-1, с. 280-283.
Миллер М.А. Археология в СССР. Издательство: Мюнхен, 1954, с. 161.
Мошкова М.Г. Археологические памятники южноуральских степей второй половины II-IV в. н. э.: позднесарматская или гунно-сарматская культура (погребальный обряд) // Российская археология, 2007, № 3, с. 103-111.
Носовский Г.В., Фоменко А.Т. Империя. АСТ. 2012.
Трепавлов В.В. История Ногайской Орды. 2-е изд., испр. и доп. Казань: Издательский дом «Казанская недвижимость», 2016, 764 с.
Тюрин А.М. Практика радиоуглеродного датирования. Часть 3. Калибровочная кривая // Электронный сборник статей «Новая Хронология», 2005, Вып. 3. [Новая хронология]
Тюрин А.М. Интеграция скифской археологической культуры в Новую Хронологию Фоменко и Носовского // Электронный сборник статей «Новая Хронология», 2007, Вып. 6 [Новая хронология]
Тюрин А.М. Имеются ли генетические следы монгольских завоеваний 13 века? // Электронный сборник статей «Новая Хронология». Вып. 10, 2010. [Новая хронология]
[Тюрин, 2017, Калмыки] Тюрин А.М. Калмыки, караногайцы, кубанские ногайцы и крымские татары – геногеографический и геногенеалогический аспекты // Журнал фронтирных исследований, 2017, № 2, с. 7-29.
[Тюрин, 2017, Литовские татары] Тюрин А.М. Генетический портрет литовских татар и феномен «Монгольские завоевания 13 века» // Вестник Оренбургского государственного университета, 2017, № 5, с. 78-82.
[Тюрин, 2017, Казахские торе] Тюрин А.М. Казахские торе не являются потомками Чингисхана // Цивилизация знаний: российские реалии: труды Восемнадцатой Международной научной конференции, Москва, 21–22 апреля 2017 г. – М.: РосНОУ, 2017, с. 93-94.
[Тюрин, 2017, Ногаи, генетика] Тюрин А.М. Калмыки, караногайцы, кубанские ногайцы и крымские татары – геногеографический и геногенеалогический аспекты // Журнал фронтирных исследований, 2017, № 2, с. 7-29.
[Тюрин, 2017, Монголоидность, хазары] Тюрин А.М. Датирование хазар по монголоидности // Астраханские Петровские чтения, 2017, с. 130-134.
[Тюрин, 2018, Ногаи, антропология] Тюрин А.М. Ногаи: антропологический аспект // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2018, Монголоидность, бронзовый] Тюрин А.М. Датирование по монголоидности популяций, относимых к бронзовому веку // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2018, Монголоидность, сарматы] Тюрин А.М. Датирование сарматов по монголоидности // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2018, Готы] Тюрин А.М. Готы: ДНК-генеалогический и антропологический аспекты // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2019, Караногайцы] Тюрин А.М. Элементы этногенеза караногайцев по данным популяционной генетики // Перекрёстки истории. Актуальные проблемы исторической науки, 2019, с. 56-60.
[Тюрин, 2019, Кубанские ногайцы] Тюрин А.М. Элементы этногенеза кубанских ногайцев по данным популяционной генетики // Перекрёстки истории. Актуальные проблемы исторической науки, 2019, с. 60-64.
[Тюрин, 2019, Элементы этногенеза] Тюрин А.М. Элементы этногенеза караногайцев и кубанских ногайцев по данным популяционной генетики // Ногайцы: XXI век. История. Язык. Культура. От истоков – к грядущему, 2019 (в печати).
[Тюрин, 2019, Ногаи] Тюрин А.М. Ногаи большой орды: путь от Приуралья до Стамбула (по данным популяционной генетики) // Ногайцы: XXI век. История. Язык. Культура. От истоков – к грядущему, 2019 (в печати).
[Тюрин, 2019, Башкиры-1] Тюрин А.М. К вопросу о монголоидных компонентах у башкир // Электронный сборник статей «Новая Хронология», 2018, Вып. 15. [Новая хронология]
[Тюрин, 2019, Башкиры-2] Тюрин А.М. К вопросу о монголоидных компонентах у башкир // XIII Конгресс антропологов и этнологов России: сб. материалов. Казань, 2–6 июля 2019 г. / Отв. ред.: М.Ю. Мартынова. – Москва; Казань: ИЭА РАН, КФУ, Институт истории им. Ш. Марджани АН РТ, 2019, с. 95.
Учанева Е.Н., Казарницкий А.А., Громов А.В., Лазаретова Н.И. Население Минусинской котловины в раннем железном веке: к вопросу о внутригрупповой и межгрупповой изменчивости // Вестник археологии, антропологии и этнографии, 2017, № 1 (36), с. 78-87.
Харузин А.И. Курганы Букеевской степи. Левенсон и К°. 1890. 118 с.
Ходжайов Т.К., Ходжайова Г.К. К проблеме формирования антропологического состава киргизов // Вестник археологии, антропологии и этнографии, 2015, № 3 (30), с. 82-91.
Balinova N., Post H., Kushniarevich A., Flores R., Karmin M., Sahakyan H., Reidla M., Metspalu En., Litvinov S., Dzhaubermezov Murat., Akhmetova V., Khusainova R., Endicott Ph., Khusnutdinova E., Orlova K., Bakaeva E., Khomyakova I., Spitsina N., Zinchenko R., Villems R. and Rootsi S. Y-chromosomal analysis of clan structure of Kalmyks, the only European Mongol people, and their relationship to Oirat-Mongols of Inner Asia. European Journal of Human Genetics,volume 27, pages1466–1474 (2019)
Damgaard P.B., Marchi N., Rasmussen S., Peyrot M., Renaud G., Korneliussen T., Moreno-Mayar J.V., Pedersen M.W., Goldberg A., Usmanova E., et al. 137 ancient human genomes from across the Eurasian steppes. Nature 557, 2018, 369–374.
Distribution of European Y-chromosome DNA (Y-DNA) haplogroups by country in percentage.
Järve M., Saag L., Scheib C.L., Pathak A.K., Montinaro F., Pagani L., Flores R., Guellil M., Saag L., Tambets K., Kushniarevich A., Solnik A., Varul L., Zadnikov S., Petrauskas O., Avramenko M., Magomedov B., Didenko S., Villems R. Shifts in the Genetic Landscape of the Western Eurasian Steppe Associated with the Beginning and End of the Scythian Dominance // Current Biology. July 2019.
Krzewinska M., Kilinc G.M., Juras A., Koptekin D, Chylenski M., Nikitin A.G., Shcherbakov N., Shuteleva I., Leonova T., Kraeva L., et al. (2018). Ancient genomes suggest the eastern Pontic-Caspian steppe as the source of western Iron Age nomads. Sci. Adv. 4, eaat4457.
Unterlander M., Palstra, F., Lazaridis, I., Pilipenko, A., Hofmanova´ , Z., Groß, M., Sell, C., Blo¨ cher, J., Kirsanow, K., Rohland, N., et al. (2017). Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat. Commun. 8, 14615.
Yunusbayev B., Metspalu M., Metspalu E., Valeev A., Litvinov S., Valiev R., Akhmetova V., Balanovska E., Balanovsky O., Turdikulova S., Dalimova D., Nymadawa P., Bahmanimehr A., Sahakyan H., Tambets K., Fedorova S., Barashkov N., Khidiyatova I., Mihailov E., Khusainova R., Damba L., Derenko M., Malyarchuk B., Osipova L., Voevoda M., Yepiskoposyan L., Kivisild T., Khusnutdinova E., Villems R. Thegenetic legacy of the expansion of Turkic-speaking nomads across Eurasia. // PLoSGenet. – 2015. V.11(4):e1005068. Impactfactor (ThomsonReuters) - 7,528. doi: 10.1371/journal.pgen.1005068
статья получена 3.10.2019
