Биологические маркеры Руси-орды и скифская археологическая культура
Тюрин Анатолий Матвеевич, к.г.-м.н.
Ключевые слова: Новая Хронология, скифы, антропология, генетика.
Оглавление
1. Постановка задачи
3. Антропологический портрет древних популяций Минусинской котловины
4. Антропологический и генетический портреты древних и современных популяций Алтая
5. Антропологический портрет древних и современных популяций Западной Сибири
6. Генетический портрет современных популяций Восточной Европы
7. Результаты сопоставления антропологического портрета скифов и популяции кобанской культуры
8. К проблеме происхождения коренных жителей Америки
9. Элементы реконструкции распространения генетических линий HG16 и Х-мтДНК
1. Постановка задачи
Общая постановка задачи - интеграция скифской археологической культуры в Новую Хронологию А.Т. Фоменко и Г.В. Носовского (НХ ФиН) [Сайт Новая Хронология], сформулирована в статье [Тюрин, 2007, Интеграция, Скифы]. В ней же приведены результаты ее решения. Биологический аспект этой задачи рассмотрен в настоящей статье. Его посылкой является следующая гипотеза: скифская археологическая культура сформирована вполне определенными способами по материальному наследию, оставленному Ордой, являющейся военной составляющей империи Руси-орды. Все необходимые сведения по этому вопросу приведены в статье [Тюрин, 2007, Интеграция, Скифы]. Орда имела вполне определенные биологические маркеры. Их можно обозначить по НХ-реконструкциям прошлого Человечества или выделить по результатам специального анализа имеющихся данных по биологическим особенностям древних и современных популяций степной и лесостепной зон Евразии. Можно определить географию их распространения и сопоставить ее с географией распространения скифской археологической культуры. Результаты сопоставления как раз и определят степень соответствия вышеприведенной гипотезы реальному положению дел. Но так получилось, что, начав анализ имеющейся информации, мы вышли за рамки решаемой задачи. Это и обусловило необходимость изложения его результатов в виде самостоятельной публикации, которая в информационном плане является частью статьи [Тюрин, 2007, Интеграция, Скифы].
2. Общие сведения
Автором публикации (Gryaznov, 1983) [Alekseev, 2001, Radiocarbon] выделено 11 географических зон распространения скифской археологической культуры: 1) север Причерноморского региона; 2) Северный Кавказ; 3) регион, прилегающий к Аральскому морю; 4) Центральный и Северный Казахстан; 5) Семиречье, Тянь-Шань и Памир; 6) степная и лесостепная зоны, прилегающие с севера и востока к Алтайским горам; 7) Алтайские горы; 8) Южная Сибирь; 9) Тува; 10) Монголия; 11) Ордос. Отметим, что в опубликованной литературе «по скифам» территорию Тувы обычно называют Центральной Азией. Скифская культура Южной Сибири локализована, главным образом, в Минусинской котловине и имеет региональное название «тагарская». Авторами публикации [Alekseev, 2001, Radiocarbon] к скифской отнесены археологические культуры Нижней Волги и Южного Урала. Ранее они относились к сарматской культуре и культурам народов Южного Урала. Однако, по ряду признаков, эти культуры являются типично скифскими.
Мы будем различать три группы биологических маркеров: антропологические, генетические и собственно биологические. Антропологические маркеры – это, прежде всего, облик представителей конкретных популяций, который определяется, главным образом, по черепам. Генетические маркеры это гены, которые не влияют на биологические особенности людей, то есть, не определяют их «конкурентоспособность» в выживании в определенных природных и социальных условиях. Собственно биологические маркеры – это гены, которые определяют биологические особенности людей и являются объектом естественного отбора. К ним относятся и гены, которые определяют группу крови человека. Группа крови, являясь одним из факторов естественного отбора, может служить биологическим маркером популяции или социальной группы. В статье рассмотрены только антропологические и генетические маркеры.
Геном человека включает три части, которые по-разному передаются в поколениях:
- хромосомы, которые передаются потомству и от отца, и от матери;
- Y-хромосома, которая передается только по отцовской линии;
- митохондриальная ДНК (мтДНК), которая передается только по материнской линии.
«К настоящему времени установлено, что митохондриальные генофонды популяций человека представлены несколькими наборами групп из определенных типов (линий) ДНК, причем каждая из них ведет свое происхождение от единственной основательницы. Для большинства групп типов мтДНК установлена региональная специфичность. Так, в генофондах народов Европы и Западной Азии (т.е. европеоидов) присутствуют главным образом 10 основных групп мтДНК (HV*, H, V, J, T, U, K, I, W, X), в генофондах монголоидных популяций Азии — тоже 10, но других групп (A, B, C, D, E, F, G, Y, Z, M*), а у негроидов — мтДНК макрогруппы L. Заметим, что коренное население Америки характеризуется редуцированным набором групп — A, B, C, D и X. Чтобы определить, откуда и когда происходили миграции, которые привели к современному генофонду анализируемой популяции, необходимо учитывать, что при его формировании была перенесена часть генетического разнообразия из популяций-основательниц.» [Деренко, 2002]. «В противоположность мтДНК маленькая Y-хромосома предается строго от отца сыновьям, то есть по мужской линии и, изучая изменчивость этой части генома, можно получить представление о происхождении мужской половины того или иного этноса.» [Захаров И.А., 2003]. Таким образом, сегодня имеется инструмент получения достоверной информации о прошлом народов.
В соответствии с Традиционной Историей (ТИ) считается, что монголоидный компонент в антропологическом и генетическом портрете жителей Восточной Европы есть следствие прихода в регион в 13 веке войска монголов. В соответствии с НХ ФиН, так называемая монгольская империя – это Русь-орда, возникшая в 14 веке в Восточной Европе и начавшая отсюда экспансию на восток, юг, и запад. Военным инструментом экспансии являлась Орда – армия, сформированная и организованная на основе определенных принципов [ХРОН4, Глава 3]. На начальном этапе Орда формировалась за счет призыва в нее молодежи с территории Руси-орды. Со временем она превратилась в «самопроизводящую» сословную структуру. Если это так, то антропологическим и генетическим маркерами Орды будет, прежде всего, европеоидность, которая сложилась в Восточной Европе в середине 2 тысячелетия н.э. Вторым маркером Орды будет высокий уровень антропологического и генетического разнообразия входящего в нее военного сословия. Возможно выявление и ее специфических антропологических, генетических, и собственно биологических маркеров. Экспансия Руси-орды на восток в 14 веке выразилась в формировании Пегой орды, которая была расквартирована в Саяно-Алтайском регионе. Наиболее полная информация по Пегой орде приведена в статье [Тюрин, Лингвистика, Скифы].
3. Антропологический портрет древних популяций Минусинской котловины
«Первыми носителями культуры эпохи металла в Сибири принято считать представителей афанасьевской культуры. Афанасьевская культура - одно из самых загадочных явлений в древней истории Сибири. Во времена перехода от эпохи камня к эпохе металла основное население Сибири было монголоидами, в то время как носители афанасьевской культуры были людьми европеоидной расы. Родственные афанасьевцам культуры были расселены широко по лесостепной зоне Евразии.» [Сибирский энеолит]. С эпитетом «одно из самых загадочных явлений в древней истории Сибири» следует согласиться. Афанасьевская культура энеолита - середина 3 - начало 2-го тыс. до н. э., действительно имеет две особенности: ярко выраженная «локальность» распространения (Минусинская котловина и Алтай) и палеоевропеоидный тип ее носителей в окружении монголоидного населения. Антропологические характеристики популяций следующих двух археологических культур бронзового века: окуневской (первая половина 2 тысячелетия до н. э.) и андроновской (середина и вторая половина 2 тысячелетия до н. э.) в просмотренной нами литературе не упоминаются. Завершает бронзовый век карасукская культура. Рубеж между карасукской и смежной с ней тагарской культурами является и рубежом бронзового и железного веков. Две последние культуры рассмотрены в статье [Тюрин, 2007, Интеграция, Скифы]. По новым результатам радиоуглеродного датирования уточнены нижний хронологический рубеж карасукской культуры: 13-12 века до н.э., и хронологические рубежи тагарской культуры: конец 10-9 века – середина 1 или 2 веков н.э. Причем, «... происхождение тагарской культуры от позднего этапа карасукской не вызывает сомнения ...» [Вадецкая, 1986]. На основе ре-фальсификации радиоуглеродных дат [Тюрин, 2005, Алгоритмы] обоснована следующая гипотеза. Карасукская и тагарская культуры Минусинской котловины являются частью общей культуры Орды и ее компонента под названием «Енисейские кыргызы», вычлененной путем применения фильтра «скифская триада». На его выходе получена скифская культура. Затем она «разрезана» репером «завершение строительства пирамиды Хеопса – начало строительства больших курганов» на две части. Одна из них – карасукская культура, отнесена к пре-скифскому времени, начало другой – тагарской, определено как начало скифской культуры в Минусинской котловине. Нижний ре-фальсифицированный рубеж карасукской культуры - первая половина 14 века, соответствует образованию Руси-орды и ее военного подразделения Енисейские кыргызы. В середине 15 века у кыргызов произошла «смена моды» на погребение тел своих сослуживцев и они начали строить большие погребальные курганы.
«Ещё один аспект происхождения тагарской культуры связан со сходством физического типа тагарцев и афанасьевцев, которое очень трудно объяснить. По утверждению антропологов, сходство это настолько специфично по всем признакам, что вряд ли оно могло явиться результатом случайного совпадения или параллельного развития. Скорее всего, оно говорит о генетическом родстве и опосредованном происхождении носителей тагарской культуры от афанасьевской (18, с. 69). Возможно, что потомки афанасьевцев, в течение веков жившие в Саянах и Горном Алтае, вернулись в степные районы.» «Наиболее труднообъяснима «этнографическая» близость тагарцев к афанасьевцам, особенно с периода коллективных могил.» [Вадецкая, 1986]. Эти «загадки» объясняются на основе нашей гипотезы. Носители карасукской и тагарской культур воины Орды - енисейские кыргызы, пришли в Минусинскую котловину из Восточной Европы. Естественно, они были европеоидами. Естественно и то, что их антропологический облик близок к облику палеоевропеоидов афанасьевской культуры. Но здесь есть одна тонкость. Если выяснится, что скифы тагарской культуры не имеют в Азии антропологических корней, то это будет свидетельствовать против хронологических и археологических теорий, основанных на свидетельстве Геродота [Тюрин, 2007, Датирование, Скифы]: «Геродот указывает, что скифы пришли из Азии, границей которой в то время считали Дон.» [Авдусин, 1980]. Не является ли афанасьевская археологическая культура искусственно сформированным фантомом, призванным обеспечить европеоидные корни скифам Азии? Если это так, то понятен и феномен «европеоиды Минусинской котловины в окружении монголоидов» и «невнятность» описаний антропологического портрета популяций окуневской и андроновской культур. Следует отметить, что наш вывод о том, что популяция тагарской культуры является европеоидной, получен логическим путем. В соответствующей главе монографии [Вадецкая, 1986] прямо об этом не говорится. Говорится только о «необъяснимом» сходстве «физического типа тагарцев и афанасьевцев», а то, что афанасьевцы палеоевропеоид – написано во всех справочниках.
В публикации [Вадецкая, 1986] приведены и другие данные, подтверждающие нашу гипотезу. «Этническая разнородность тагарцев прослеживается в период сложения культуры и на раннем её этапе, т.е. в каменноложско-баиновское время. С середины подгорновского этапа погребения становятся единообразными.» «... тагарская культура в конечном итоге сложилась из разных локальных групп, сосуществовавших ещё в карасукскую эпоху ...». Конечно, в Минусинскую котловину пришла орда, сформированная из групп народов Восточной Европы, различающихся между собой, и антропологически, и культурно. Примерно за 100 лет совместного проживания в географически замкнутом регионе произошло формирование единой культуры.
В публикации [Вадецкая, 1986] приведены результаты датирования тагарской и таштыкской культур Южной Сибири. Радиоуглеродные даты, характеризующие тагарскую культуру попали в интервал 1-2 веков н.э., таштыкскую (6 дат) – в 1 век до н.э. – 1 век н.э. То есть, таштыкская культура является одновозрастной завершающему этапу таганской культуры. Отмечается, что «... генетическая связь татарской и таштыкской культур не вызывает сомнения, но в сложении последней немалое участие приняло новое население, пришедшее в Минусинскую котловину со своей культурой.» Ре-фальсифицированная дата «1 век до н.э.» соответствует первой половине 18 века. Что произошло в Минусинской котловине в этот период? Мы предполагаем, что енисейские кыргызы, являющиеся военным сословием одного из пост-Ордынских формирований, активно участвовали в войнах манЖуРов/манГуЛов, создавших в Китае государство ордынского типа [ХРОН5, часть 2, глава 6]. Из Тувы в Китай ушли все кыргызы вместе с частью зависимых от них социальных общностей [Боронин] (этот вопрос рассмотрен в публикации [Тюрин, Лингвистика, Скифы]). Они на свое прежнее местожительство не вернулись. Возможно, часть подразделений кыргызов Минусинской котловины, побыв в Китае, вернулась на свою родину. По их материальному наследию сформирована таштыкская археологическая культура. В связи с этим, мы прокомментируем информацию, приведенную в следующей цитате. «Что же касается таштыкских грунтовых могильников, то в них привилегированными покойниками были мужчины-воины, трупы которых подвергали кремации, но есть основания считать, что эти воины в отличие от тагарцев были монголоидами. Хотя среди мумий в могилах совсем нет, а в склепах почти нет представителей дальневосточного или центральноазиатского облика, маски, одетые па лица трупов в могилах, отражают европеоидов и монголоидов, а маски на куклах в склепах запечатлели 3 антропологических типа населения: европеоидов, монголоидов и смешанных, т.е. они демонстрируют не только распространение монголоидности, но и уже сложение, благодаря этому, нового антропологического типа населения (132, с. 315).» [Вадецкая, 1986]. Вывод о монголоидном компоненте тыштыкской популяции сделан только по погребальным маскам. Скорее всего, погребальные маски отражают «моду на монголоидность», которая появились в Китае в связи с начавшейся ассимиляцией его правящего сословия - манЖуРов/манГуЛов.
Завершают антропологический портрет древних популяций Минусинской котловины этнографические данные, приведенные в [ХРОН4, глава 3] со ссылкой на работу по истории Сибири. «В половине ДВЕНАДЦАТОГО ВЕКА в восточной и центральной Азии жили самостоятельные племена, носившие название КАЗАЧЬИХ ОРД. Наиболее значительная «казачья орда» жила в верховьях реки Енисея и занимала земли на востоке от озера Байкал и на западе до реки Ангара. В китайских хрониках орда эта называлась хакассы, что по исследованию европейских ученых равнозначно слову «казак». По запискам, оставленным современниками, - «хакасы» или «казаки» принадлежали к индо-иранской расе. Они были белокуры и светловолосы; высокие ростом, с зелено-голубыми глазами; храбры, горды и в ушах носили кольца. (Рихтер, немецкий историк 1763-1825 гг., «Иохим», Записки о Монголии).» К этому дадим два комментария. Индо-иранская раса – это и есть европеоиды, а казачья орда, проживающая в верховьях Енисея – енисейские кыргызы.
Таким образом, в Традиционной Археологии не имеется достоверных данных о генетических корнях популяций какрасукской и тагарской культур Минусинской котловины, являющихся европеоидными. Не имеется и гипотез, обоснованных фактическими данными и объясняющих появление европеоидов среди монголоидного населения. Эти «не имеется» маскируются невнятным изложением антропологического вопроса при описании археологических культур региона. Зато, европеоидность популяций какрасукской и тагарской культур, а конкретно, енисейских кыргызов, подтверждает две взаимосвязанные гипотезы: енисейские кыргызы – воины Руси-орды 14-16 веков и пост-Ордынских формирований 17-18 веков, предки которых пришли в Южную Сибирь в 14 веке из Восточной Европы; карасукская и тагарская культуры сформированы по материальному наследию енисейских кыргызов.
4. Антропологический и генетический портреты древних и современных популяций Алтая
Хорошая сохранность скифских захоронений Алтая (пазырыкская культура) дала возможность выполнить палеоантропологические и палеоэтногеномические исследования останков людей. Эти данные прямо характеризуют популяцию, соответствующую скифской археологической культуре. В публикации [Voevoda] обобщены итоги палеоантропологических (краниологических) исследований популяций пазырыкской культуры. Установлено, что они имели монголоидные и европеоидные компоненты. Монголоидные компоненты включают два антропологических типа. «... автохтонный, был найден в Алтайском крае на границе Неолита и Энеолита в людях, погребенных в Нижнее-Титкескенской и Каменной пещерах, и во второй половине II тысячелетия до н.э. в населении каракольской культуры. Типичная комбинация антропологических характеристик параметров этого типа ... встречается в настоящее время в казахах, киргизах и некоторых группах хакасов и Южно-Алтайских народов. Второй тип, палеосибирский, доминировал на территории около озера Байкал в течение Неолита. В настоящее время, он встречается только у эвенков северобайкальского региона.» Отмечено отсутствие абсолютного параллелизма в антропологических и молекулярно-генетических характеристиках популяций пазырыкской культуры. Результаты сопоставления палеоантропологических исследований популяций пазырыкской культуры и антропологических исследований современных народов Саяно-Алтайского и сопредельных регионов рассмотрены в публикации [Tur, 2003]. Последние имеют европеоидные и монголоидные компоненты. Отмечено, что древние пазырыкцы имели более выраженные европеоидные компоненты, чем представители современной южно-сибирской расы. В народах Северного Алтая доминирует европеоидный компонент, в народах Южного Алтая – монголоидный. Сделан вывод: типичные антропологические характеристики народов Северного Алтая могут рассматриваться как современные модификации характеристик популяций пазырыкской культуры. Наиболее четко это просматривается для кумандийцев, телеутов, тубаларов и горных шорцев. В этот же кластер попали барабинские татары и одна из групп башкир (Mavlutovo Bashkirs). Телеуты – это одно из самоназваний воинов соединения Теле, правого крыла Пегой орды [Тюрин, 2007, Лингвистика, Скифы]. Предполагается, что скифские курганы высокогорного алтайского плато Укок – это захоронения вождей и воинов Пегой орды. Следовательно, сегодняшние телеуты как раз и должны быть антропологически близки популяции пазырыкской культуры. Кумандийцы и тубалары – малые народы Алтая. Про них мы ничего сказать не можем. Можно предположить, что барабинские татары и Mavlutovo Bashkirs – потомки популяции пазырыкской культуры. Шорцы рассмотрены ниже. Одним из показаний расовой принадлежности людей является цвет их волос. Сетлые волосы – индикатор европеоидности «У мужчины с плато Укок были светлые рыжеватые волосы, подстриженные спереди и лежащие на лбу в виде длинной челки, на макушке оставлен большой участок не остриженных волос, заплетенных в две косы, каждая из двух прядей, спускающихся ниже плеч.» [Полосьмах].
В публикации [Тур, 2004] приведен еще один блок краниологических данных по пазырыкской культуре. «Опубликованы лишь средние размеры суммарной краниологической серии из рядовых погребений пазырыкской культуры, в состав которой помимо черепов из Кызыл-Джара были включены также черепа из могильников Талдура I, Кара-Коба II, Кызык-Телань I и Нижний Тюмечин (Дремов В.А., 1990, с.132-142). Опираясь на обобщенные характеристики этой серии, автор исследования пришел к заключению, что рядовое население пазырыкской культуры в своей основе было европеоидным и в отличие от пазырыкской знати имело в своем составе лишь небольшую долю монголоидной примеси центрально-азиатского происхождения. Было установлено также, что ближе всего к рядовым пазырыкцам синхронное население Тянь-Шаня и Тывы (Дремов В.А., 1990, с.134-136).» То есть, европеоидность рядового населения пазырыкской культуры не вызывает сомнений. Кроме того, отмечена сходная с ним европеоидность синхронного населения Тянь-Шаня и Тувы. А это две зоны распространения скифской культуры. Здесь следует иметь в виду, что вывод о европеоидности сформирован по результатам изучения захоронений и, следовательно, прямо не распространяется на все население этих регионов. Но он прямо распространяется на все скифские популяции, курганы и захоронения которых находятся в Центральной Азии (Тува), Алтае и Тянь-Шане. Повышенный компонент монголоидности пазырыкской знати - военных руководителей Пегой орды, объясняется очень просто. Военные руководители были обязаны по долгу службы брать себе в жены дочерей вождей монголоидных племен, проживающих на территории Саяно-Алтайского региона и в сопредельных областях. И последнее. Урочище Кызыл-ДЖАр это обычный красный Яр, где ЯР – один из самых распространенных топонимов (как правило, в сочетании с другими словами) на территории Орды.
В таблице 1 приведены данные по распространенности различных идентифицированных типов мтДНК (%) среди народов Саяно-Алтайского региона и сопредельных областей [Захаров И.А., 2003]. Выше мы установили европеоидность енисейских кыргызов по антропологическим и этнографическим данным. Их потомки, хакасы, являются по типам мтДНК европеоидами на 18,6%. Шорцы живут в основном в Кузнецкой области, Хакасии и на Алтае. «По антропологической классификации шорцев принято относить к уральскому типу большой монголоидной расы. В то же время по ряду морфологических и краниологических признаков шорцы выходят за рамки как уральского, так и южносибирского антропологических типов.» [Шорцы. http://russia.rin.ru/guides/4727.html]. Эта особенность шорцев объясняется очень просто. «В 17-18 веках с Шорцами слились пришедшие с Севера (Иртыш, Барабинская и Кулундинская степи) кочевники-скотоводы телеуты.» [Шорцы. http://www.narodru.ru/peoples1301.html]. Скорее всего, они и обеспечили наличие у современных шорцев европеоидности (по мтДНК) на 35,3%, а также их антропологические характеристики, которые «выходят за рамки как уральского, так и южносибирского антропологических типов.» Второе самоназвание воинов Теле – телесы. Часть телесов вошла как сеоки в алтайские народы [Тюрин, Лингвистика, Скифы]. Они и обеспечили их европеоидность на 20,7%. Классические скифские курганы распространены и в Туве, а европеоидность сегодняшних тувинцев не превышает фонового уровня. Почему? Ответ прост. В начале 18 века енисейские кыргызы из Тувы ушли. Поэтому современные народы региона и не унаследовали антропологических и генетических компонентов строителей скифских курганов 14-17 веков. А антропологический облик последних был идентичен антропологическому облику скифов пазырыкской культуры [Тур, 2004].
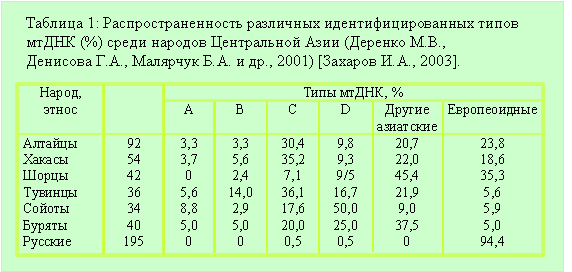
|
В публикации [Молодин, 1999] приведены обобщенные результаты палеогенетического анализ генофонда Саяно-Алтайского региона и сопредельных областей. Основу поалеогенетических данных составили археологические результаты изучения Пазырыкской культуры. Установлена генетическая близость носителей пазырыкской культуры и современных самодийцев (селькупов, кетов), сделан вывод о существовании «расогенетических связей у населения Алтая и полуоседлых племен скотоводов Парфии Маргианы, Северной Бактрии с эпохи бронзы ...».
Антропологические характеристики и классификация алтайцев, приведены и в публикации [Ялбачева]. «Большие антропологические различия между южными и северными группами алтайцев не вызывают сомнения. Южные алтайцы относятся нашими антропологами, как и тувинцы, буряты, монголы, якуты к наиболее монголоидному, так называемому центрально-азиатскому и южно-сибирскому типам. У северных алтайцев монголоидность основных антропологических признаков выражена гораздо слабее, но у них наличествуют и европеоидные признаки. Северные алтайцы относятся антропологически, вместе с рядом угорских и солиодийских народностей, к уральскому типу. Таким образом, северные алтайцы, особенно шорцы и кумандинцы, обнаруживают общность антропологического типа по измерительным и описательным признакам не с южными алтайцами, а с обскими уграми: хантами и манси.» Отмечено, что «Проблема происхождения тюркоязычных племен Алтая давно относится к числу наиболее трудных и неразработанных в этнографии Сибири.»
Антропологических и этногеномических данных вполне достаточно для вывода о том, что популяции пазырыкской культуры Алтая имела существенный европеоидный компонент. Это первый маркер Орды. Данных вполне достаточно и для вывода: народы Алтая являются потомками популяций пазырыкской культуры. В свою очередь, народы Алтая имеют большие внутрипопуляционные антропологические различия и наибольшее разнообразие мтДНК. Это второй маркер Орды. Наличие двух маркеров Орды на Алтае подтверждает гипотезу: пазырыкские курганы – это захоронения ее руководителей и воинов.
5. Антропологический портрет древних и современных популяций Западной Сибири
Палеоантропологические характеристики народов Западной Сибири и сопредельных регионов по краниологическим данным рассмотрены в публикации [Багашов, 2002]. Европеоидные компоненты в популяциях Западной Сибири, тяготеющие к югу региона, появились в эпоху бронзы. В популяциях таежной зоны преобладал монголоидный компонент. «Антропологический состав населения Западной Сибири раннего железного века становится еще более сложным. Своеобразие антропологического типа западносибирских популяций определяют несколько основных компонентов, удельный вес которых в локальных группах варьирует. В составе населения саргатской и гороховской культур основным является европеоидный расовый вариант с мезо-брахикранной формой черепа, широким невысоким и умеренно профилированным в горизонтальной плоскости лицевым скелетом. Антропологическую специфику кулайской серии (могильник Каменный Мыс) определяет монголоидный компонент, характерными чертами которого являются малая высота уплощенного лицевого скелета, слабое выступание носовых костей при средневысоком переносье. Помимо вышеназванных основных компонентов, определенную роль в расогенезе лесостепных популяций сыграл европеоидный комплекс с высоким и узким лицом, а также монголоидный с широким и высоким лицом. В отдельных выборках, в частности в Прииртышье, удалось зафиксировать примесь монголоидного компонента, обладающего долихокранной формой черепа.» «В средние века антропологический покров южной части Западной Сибири изменяется по сравнению с предшествующим временем. Основной европеоидный компонент расовой структуры саргатского населения не имеет прямых аналогий в составе средневековых популяций.» «В близкое к современности время на территории Западной Сибири фиксируется большое разнообразие антропологических типов. В некоторых случаях проявляется этногенетическая связь современных народов с европеоидным населением раннего железа. Более или менее значимая роль может быть отведена ему лишь в генезисе физического облика отдельных групп тоболо-иртышских татар.»
Одна из особенностей антропологической истории Западной Сибири описана так. «В отличие от Алтая и Хакасии, на территории Западной Сибири существовали популяции, в антропологическом типе которых фиксируется ряд особенностей, определенных не столько примесью монголоидных и/или европеоидных черт, сколько специфическим сочетанием расовых признаков, в которых нарушена нормальная историческая корреляция, что указывает на сохранение следов недифференцированности. В итоге именно этим определяется своеобразие древнего и современного населения Западной Сибири.»
По результатам реконструкции антропологической истории популяций юга Западной Сибири получилось, что в раннем железном веке в их облике имелись ярко выраженный европеоидный компонент. Но в облике популяций, населяющих регион в средневековье, европеоидный компонент популяций раннего железного века не проявился. Зато он проявился в облике современных народов. Это названо нарушением нормальной исторической корреляции расовых признаков. Эти результаты прекрасно иллюстрируют наш алгоритм формирования скифской археологической культуры. Она сформирована по материальному наследию Орды и пост-Ордынских формирований 14-18 веков, частью которого являются захоронения ее европеоидных, в своей массе, воинов, и «сдвинута» в поздний бронзовый - ранний железный века. Вот и получилось, что европеоидный субстрат искусственно вставлен в монголоидный, а позднесредневековый европеоидный субстрат связан не с раннесредневековым, но с субстратом раннего железного века. То есть, он связан сам с собой. Характерно и то, что наиболее контрастно проявилась связь «европеоидов раннего железного века» с отдельными группами тоболо-иртышских татар. Слово ТАТАР является одним из лингвистических маркеров Орды [Тюрин, 2007, Лингвистика, Скифы].
6. Генетический портрет современных популяций Восточной Европы
Этногеномика народов Восточной Европы рассмотрена в публикации [Хуснутдинова, 2003]. Проанализированы геномы 1500 индивидов из 8 популяций (башкиры, татары, русские, чуваши, удмурты, коми, мордва, марийцы). «Анализ европеоидного и монголоидного вклада в материнские генетические линии народов Волго-Уральского региона не выявил корреляции языка и геномного состава этнических групп. На языках тюркской группы, привнесенных из Азии, говорят не только башкиры (65% монголоидности), но и татары и чуваши, у которых преобладает европеоидный генетический компонент. В остальных популяциях региона вклад монголоидного компонента составляет от 12% у русских до 20% у удмуртов. Русские, проживающие на территории этого региона, имеют 10-12% монголоидных типов мтДНК, а русские из Рязанской и Курской областей – только 2-3%. Это можно объяснить смешением русских с тюркоязычными народами на территории Волго-Уральского региона. Интересно, что некоторые материнские линии у разных народов, например, у русских, татар и марийцев, оказались общими. Это показывает глубокое родство народов, говорящих на разных языках, придерживающихся разных религий и традиций.» Особое внимание уделено рассмотрению линий 12 и 16 мужской Y-хромосомы (HG12 и HG16). «Линия 16 практически отсутствует в западноевропейских популяциях, но частота ее высокая среди народов Балтии - эстонцев и финнов, а также у народов Волго-Уральского региона, особенно удмуртов и коми-зырян. ... Для линии 16 Y-хромосомы уровень генетического разнообразия намного выше в популяциях Восточной Европы (чуваши, татары), чем в изученных популяциях Сибири. Хотя удмурты и имеют очень высокую частоту линий 12 и 16, уровень генетического разнообразия у них невысокий по сравнению с другими популяциями Европы. Данные о низком уровне генетического разнообразия удмуртов получены и по материнской линии при анализе полиморфизма митохондриальной ДНК. Все это свидетельствует о несомненной роли эффекта основателя и дрейфа генов в демографической истории удмуртов. Анализ распространения и разнообразия линии 16 Y-хромосомы среди восточноевропейских популяций показывает то, что местом ее «рождения», возможно, является Восточно-Европейская равнина. Согласно филогеографическому анализу этой линии в Евразии, она начала распространяться с запада на восток. В то же время частота распространения линии 12 - предковой для линии 16 - меньше, чем для линии 16». Предположение о том, что местом «рождения» линии HG16 является Восточно-Европейская равнина, заслуживает самого серьезного внимания в контексте выявления новых генетических маркеров Орды.
7. Результаты сопоставления антропологического портрета скифов и популяции кобанской культуры
Автор публикации [Тихонов] выполнил сопоставление краниологических и остеологических характеристик черепов могильника кобанской культуры Уллубаганалы (7-6 века до н.э.) расположенного в 30 километрах к западу от Кисловодска и скифов Северного Кавказа, Причерноморья и Приднепровья. По его результатам сделаны следующие выводы. «Мы отметили принадлежность серии из могильника Уллубаганалы к северокавказскому кругу форм, для которого характерны долихокрания, узкое сильнопрофилированное лицо, крупный выступающий нос. Вместе с тем обнаружилось высокое сходство нашей серии со скифскими группами Приднепровья и Причерноморья, причем сходство, превосходящее сходство с северокавказскими группами. Возможных объяснений этому может быть два: либо антропологическое взаимодействие между скифскими и кобанскими племенами было до такой степени тесным, что первоначальный антропологический облик населения кобанской культуры просто растворился в массе скифских племен, либо существовал единый антропологический пласт, присущий как кочевому населению степи, так и оседлому населению Северного Кавказа. Первая возможность представляется нам маловероятной антропологически и слабо аргументированной исторически. Действительно, по данным археологии и свидетельствам письменных источников скифское влияние на Северном Кавказе сводилось к спорадическим проявлениям военной активности (скифы появлялись на Северном Кавказе в ходе своих военных походов на Ближний Восток). Естественно, что в подобных условиях, ни о каком активном обмене генами, путем брачных связей, не может быть и речи (Крупнов, 1960). Поэтому мы склонны считать, что в скифское время существовал единый антропологический пласт, отличительными чертами которого были долихокрания, высокий череп, узко резкопрофилированное лицо, крупный выступающий нос.» К этому можно добавить, что «скифское время» в антропологическом плане - это период с 14 века н.э. до сегодняшнего дня. «Единый антропологический пласт» - это единая популяция западной части Орды сложившаяся, в целом, в 14 веке. По ее материальному наследию сформированы скифская, кобанская и другие археологические культуры.
8. К проблеме происхождения коренных жителей Америки
Фактические данные, полученные в рамках программы поиска в Центральной
и Северной Азии народов, родственных коренным американцам (америндам),
и результаты их интерпретации приведены в публикации [Захаров И.А.,
2003]. Решается эта задача на основе предельно простой посылки.
Требуется найти популяции, в генофонде которых имеются все 5 типов
линий мтДНК, редуцированного набора америндов. Причем эти популяции
и америнды должны иметь и сходные Y-хромосомы. Очертив сегодняшний
ареал проживания ближайших родственников америндов на территории
Центральной и Северной Азии можно определить место, где проживала
популяция прародительница последних и реконструировать путь ее миграции
в Америку. Фактические данные, полученные по результатам исследований,
приведены в таблицах 1 и 2.
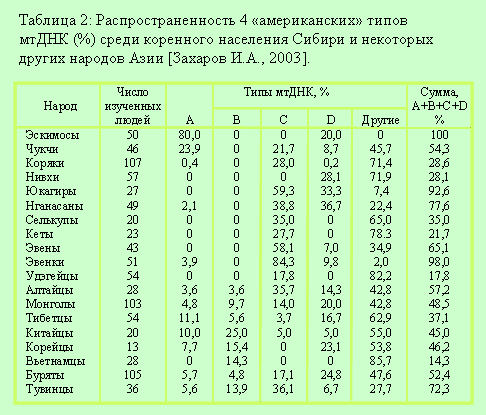
По результатам анализа фактических данных, приведенных в таблицах 1 и 2 можно сделать однозначный вывод: родственными народами америндов являются алтайцы, монголы, тибетцы, китайцы, буряты, тувинцы, хакасы и сойоты. Это без учета Х-мтДНК, которая имеется у североамериканских индейцев якима, навахо, нуу-чах-нулс, оджибва (до 30%). У европеоидов Х-мтДНК распределена мозаично (с невысокими частотами) у населения Европы, Кавказа, Западной Азии [Деренко, 2002], и до момента начала исследований (конец 90-х годов 20 века) ее наличие у сибирских народов не отмечалось. По результатам исследований [Захаров И.А., 2003] она выявлена у алтайцев. Следовательно, алтайцы являются единственным родственным народом америндов. Но имеется одна проблема: 23,8% мтДНК алтайцев относится к европеоидным линиям. Следовательно, их Х-мтДНК имеет, скорее всего, европеоидное происхождение. То есть, для признания алтайцев родственным народом америндов надо доказать, что их Х-мтДНК не имеет европеоидное происхождение. «Компьютерный анализ данной последовательности ДНК показал, что это не поздняя европейская примесь - алтайский вариант X оказался древним, который может быть сближен с предковой формой вариантов типа X современных европейцев и америндов.» [Захаров И.А., 2003]. Процитированную фразу нужно понимать так: Х-мтДНК не привнесена на Алтай переселенцами из Восточной Европы (прежде всего русскими), которые впервые появились в этом регионе в первой половине 18 века. Из этого вовсе не следует, что Х-мтДНК алтайцев является древней. Из этого следует только то, что Х-мтДНК попала на Алтай раньше, чем там появились русские переселенцы. Воины орд, сформированных в Восточной Европе, пришли в Саяно-Алтайский регион в начале 14 века. Они и принесли сюда Х-мтДНК. Этот же вывод можно сделать и по рисунку из публикации [Деренко, 2003]. Имелся единый предковый тип Х-мтДНК для алтайцев, европеоидов и америндов, а Х-мтДНК алтайцев не «занимают промежуточное положение между европейскими и североамериканскими» и это не «свидетельствует о заселении Америки из Центральной (или Южной) Сибири». Вообще, попытка «породнить» алтайскую Х-мтДНК с америндовской в обход европеоидов не только бессмысленна, но и некорректна. Это тот случай, когда решение одной проблемы порождает другую проблему. В случае признания того, что алтайская и европеоидная Х-мтДНК являются существенно отличными линиями, придется искать общего предка алтайцев и европеоидов, носителя предкового типа Х-мтДНК, причем он не должен иметь других азиатских и европеоидных линий мтДНК. В противном случае не удастся объяснить, почему от него к европеоидам не попали азиатские линии, а к предкам алтайцев, мигрировавших в Америку – европеоидные. Другими словами, он должен иметь всего одну мтДНК - предковую Х-мтДНК.
Тем не менее, родственные народы (по материнской линии) америндов определены. Это народы, проживающие сегодня на Юге Сибири и в Центральной Азии и имеющие 4 основные «американские» мтДНК. Логично предположить, что эти же народы являются родственными америндов и по отцовской линии. Однако здесь автор публикации [Захаров И.А., 2003] невнятно объяснил, что «Имея это в виду, мы начали изучать изменчивость и Y-хромосомы у народов Центральной Азии. Был получен и частично опубликован значительный материал, который, однако, не позволяет пока нам сделать определенные выводы.» То есть, полученные данные не только не позволяют сделать определенные выводы, но и полностью не опубликованы. Наш вывод вполне определенный. Конкретные народы, для которых сделан вывод об их родстве с америндами по материнской линии, не являются их родственниками по отцовской линии. Это следует и из результатов исследований, приведенных в публикации (Santos F.R., Pandya A., Tyler-Smith C. et al., 1999) [Захаров И.А., 2003]. «Они изучили изменчивость Y-хромосомы у ряда сибирских народов, в том числе бурятов, алтайцев, кетов, в сравнении с народами Северной и Южной Америки и других континентов. Оказалось, что наибольшее сходство с америндами показывают кеты - маленький народ, ныне живущий вдоль среднего течения р. Енисей, но происходящий, как предполагают антропологи, из Алтае-Саянского нагорья. Высокая частота одного из характерных для америндов варианта Y- хромосомы обнаруживается и у бурят. В результате проведенных исследований был сделан вывод (Karafet T.M., Zegura S.L., Posukh O. et al.), что территорией, откуда началась миграция азиатских предков америндов, была область от Алтая на западе до Забайкалья на востоке, включающая Саяны и северные районы Монголии.» Здесь произошла подмена смысла поставленной задачи. На первом этапе ее решения – поиск народов родственных америндам, на втором – локализация территории их проживания. Вместо родственных народов обозначена географическая область, генетические характеристики народов которой в совокупности позволяют сконструировать генетический портрет америндов.
Таким образом, прародина америндов локализована и намечен путь миграции их предков в Америку. Начальный пункт этого пути – область от Алтая до Забайкалья, промежуточная – Берингов пролив, конечная – просторы Северной и Южной Америки. Но здесь имеется еще одна проблема. Прошедшие этим путем предки америндов должны были иметь строго 5 фиксированных линий мтДНК, это во-первых, и во-вторых, оставить свои генетические следы на всем его протяжении. А последних как раз и нет. Народы, проживающие на просторах Севера Азии, Дальнего Востока, и Севера Америки не имеют всего набора «американских» линий мтДНК (смотри таблицу 2). Столь подробный разбор фактических данных и объединяющей их гипотезы о родственных народах америндов понадобился нам для того, чтобы показать, что рассуждения о не европеоидном источнике Х-мтДНК алтайцев и, следовательно, америндов являются надуманными. Х-мтДНК алтайцев – часть их европеоидных линий, общее количество которых составляет 23,8%. Х-мтДНК америндов - это другая проблема, детальное рассматрение которой следует выполнить в рамках НХ-реконструкций возникновения американских «доколумбовских» цивилизаций.
В публикации [Деренко, 2002] приведены результаты исследования структуры и разнообразия генофондов современных 12 популяций коренного североазиатского населения. Суммарная выборка составила 936 человек из различных лингвистических групп алтайской и палеоазиатской языковых семей. Основу выборки составили тюркоязычные народы Саяно-алтайского региона: алтайцы, хакасы, шорцы, тувинцы, восточные тувинцы (тоджинцы), тофалары и сойоты. Эти данные частично рассмотрены в публикации [Захаров И.А., 2003]. По материнской линии митохондриальные генофонды сибирских народов характеризуется высоким разнообразием, которое определяется разным соотношением монголоидных и европеоидных линий мтДНК. Частота последних увеличивается в западном направлении, достигая максимума (35.7%) у шорцев и алтайцев. Подробно рассмотрена проблема линии X-мтДНК (мы эту проблему уже рассмотрели). Наиболее интересными являются HG12 и HG16 (HG12 предковая по отношению к HG16) линии мужской Y-хромосомы. Их нельзя однозначно отнести к числу монголоидных или европеоидных. Отмечено, что имеются проблемы с трактовкой возникновения этих линий и их генетического вклада в происхождение народов Северной Евразии. По результатам исследований сделан вполне определенный частный вывод об особенностях заселения Америки народами Северной Азии. «Таким образом, в настоящее время с достаточной долей уверенности можно считать, что палеопопуляции Южной и Восточной Сибири были источником нескольких (по меньшей мере — двух) миграций в Америку. В ходе одной из миграций — алтае-саянской — в Америку могли попасть, среди прочих типов Y-хромосом и мтДНК, палеоевропеоидные мужские линии группы HG1, на основе которой сформировался чисто «американский» тип Y-хромосомы — группа HG18. В период другой волны расселения, шедшего из Байкальского региона, по всей видимости, распространилась группа HG10.». Сделано и частное предположение: возможно, народы Саяно-Алтайского региона участвовали в этногенезе саамов.
На первый взгляд способ выявления родственных америндам популяций Северной и Центральной Азии, основанный на анализе соответствия их генофондов, является вполне корректным. Но это не так. При его применении будет автоматически обеспечено отнесение народов, имеющих наибольшее разнообразие типов мтДНК, в родственные америндам. Собственно говоря, так и получилось. В число родственных им народов попали алтайцы, имеющие большие внутрипопуляционные антропологические различия и наибольшее среди популяций региона разнообразие мтДНК. А вот такой народ как эскимосы, имеющие всего две линии мтДНК – А и D, в родственные народы америндам по рассматриваемому формальному признаку не попадают. Эта ложная посылка (поиск родственных народов по формальному соответствию их генофондов или антропологических компонентов) является основой феномена «Алтай и сопредельные регионы - предполагаемое место формирования общностей народов: америндов, скифов, сарматов, гуннов, тюрков». Большие внутрипопуляционные антропологические различия и наибольшее разнообразие мтДНК алтайцев говорит только о том, что в этом регионе произошло смешение многих монголоидных и европеоидных народов. То есть, многие народы или сформированные на их основе социальные общности пришли в этот регион и жили здесь. Но по наблюдаемому сегодня разнообразию не представляется возможным сделать какие-либо выводы о том, что народы с редуцированным относительно алтайцев генофондом, сформировались на Алтае. Они могли сформироваться в любом регионе Евразии или даже за ее пределами, а затем принести свои гены на Алтай.
Общий вывод однозначен: Имеются серьезные проблемы палеоэтногеномических реконструкций народов Алтая и Америки. Решение этих проблем в рамках доминирующих археологических и антропологических теорий порождает другие проблемы. Это указывает на то, что доминирующие археологические и антропологические теории не соответствуют реальности.
9. Элементы реконструкции распространения генетических линий HG16 и Х-мтДНК
Автор публикации [Хуснутдинова, 2003] сделал предположение: местом «рождения» линии HG16 является Восточно-Европейская равнина. Можно предположить, что здесь же «родилась» линия Х-мтДНК. Гипотеза миграции этих линий за пределы региона выглядит следующим образом. Отцовская линия HG16 распространилась практически в пределах всей Северной Азии, а материнская линия X-мтДНК смогла достичь только Саяно-Алтайского региона, где ее носителями сегодня являются алтайцы. То есть, представители восточно-европейских народов как полноценные социальные общности, включающие мужчин и женщин, при миграции на восток не продвинулись дальше Алтая. А в миграции в регионы, расположенные восточнее и севернее Алтая, принимали участие только мужчины восточно-европейских народов, или их социальных общностей, достигших Алтая. В недалеком прошлом отдельные группы полноценных социальных общностей из Восточно-Европейской равнины мигрировали в Западную Европу и Западную Азию, а так же на Кавказ, сформировав в них локальные области сегодняшнего распространения линии Х-мтДНК. Предположения о том, что местом «рождения» линий HG16 и X-мтДНК является Восточно-Европейская равнина, сделаны только по этногеномическим данным без учета какой-либо априорной информации. Это относится и к нашей гипотезе их миграции за ее пределы. Гипотезу рекомендуется принять во внимание при выполнении НХ-реконструкций истории народов Евразии.
10. Выводы и рекомендации
Частные выводы сделаны в разделах статьи. Здесь же приведен общий вывод по результатам решения поставленной задачи и даны рекомендации по развитию наиболее перспективных направлений поиска по теме «Антропология, генетика и НХ ФиН».
1. Выделено два маркера Орды: европеоидность и высокий уровень разнообразия. Эти маркеры однотипны для антропологических и генетических характеристик популяций. Предполагается, что Орда имеет еще два специфических генетических маркера: Х-мтДНК (передается только по материнской линии) и HG16 (передается только по отцовской линии). Представляется целесообразным выполнить целенаправленный анализ распределения частот этих линий в Евразии и Африке с построением гипотезы их распространения на основе НХ-реконструкций истории Человечества, выполненных в рамках НХ ФиН. Скорее всего, удаться выявить и другие генетические маркеры Орды. Информационная база для их поиска имеется.
2. В пределах зон распространения скифской археологической культуры, находящихся в регионе проживания монголоидного населения (Южная Сибирь, Центральная Азия, Алтай, степная и лесостепная зоны, прилегающие с севера и востока к Алтайским горам, юг Западной Сибири) имеются антропологические и генетические маркеры Орды. Они имеются как у популяций, материальное наследие которых включено в скифскую культуру, так и у народов и социальных общностей, сегодня проживающих в этих регионах. Это является антропологическим и генетическим подтверждением гипотезы о том, что скифская археологическая культура сформирована вполне определенными способами по материальному наследию, оставленному Ордой, являющейся военной составляющей империи Руси-орды.
3. В НХ ФиН обоснована гипотеза о том, что в создании «доколумбовских» цивилизаций Америки в 15 веке принимали участи выходцы из Руси-орды и Западной Европы [ХРОН6, глава 14]. Если это так, то в популяциях америндов должны быть их генетические следы. Можно ставить задачу их выявления.
11. Послесловие
В 1993 году археологический отряд Института археологии и этнографии Сибирского отделения РАН при изучении скифских монументов алтайского плато Укок нашел скифскую мумию, принадлежащую к Пазырыкской культуре. Далеко не первую по счету и не последнюю. Эта мумия получила известность как «Пазырыкская принцесса» или «принцесса Кадын». Мы не будем рассматривать здесь все аспекты грязной истории, связанной с этой мумией. Приведем только одну цитату и наши комментарии к ней. «Тем не менее, местные шаманы продолжают утверждать, что и место, и мумии являются священными для алтайцев. Заявляется также, что мумия принадлежит принцессе Кыдын (Кадын), которая во все времена чтилась алтайцами. Однако ученые с помощью генетических исследований установили, что найденные мумии относились к скифам, а не тюркам и, следовательно, быть при жизни тюркской принцессой и принцем не могли.» [Леонид Драчевский]. Ученые, на которых ссылаются авторы публикации, обманывают общественность. По результатам антропологических и генетических исследований можно почти наверняка утверждать, что потомками популяции пазырыкской культуры являются народы Северного Алтая. Нет ничего удивительного, что их шаманы считают плато Укок священным для алтайцев местом. Погребение «Пазырыкской принцессы» состоялось, скорее всего, в 17 веке, то есть в пределах фольклорной памяти алтайцев. А еще ученые обманывают общественность в том, что по результатам генетических исследований можно установить принадлежность индивида к тюркам, или доказать обратное. Изощренность этого обмана заключается в том, что тюрки – это антропологически и генетически разные народы, говорящие на тюркских языках.
Источники информации
Авдусин Д.А., Полевая археология СССР,
М., 1980.
http://www.archeologia.ru/Library/Book/6420eba768ac Портал: Археология России.
http://www.archeologia.ru/
Багашов А. Н. Сложение уральских народов в свете антропологических данных. 2002 Институт Истории и Археологии УрО РАН.
http://northcongress.ural.ru/index/ru/arh/public?r_id=140 Сайт Северный Археологический Конгресс. http://northcongress.ural.ru/
Боронин О.В. «Снятие» проблемы двоеданничества и двоеподданства
коренного населения бассейна Верхнего Енисея в первой половине XVIII в.
http://new.hist.asu.ru/biblio/borod3/403-407.html#text13 Интернет-портал по истории Алтая http://new.hist.asu.ru/
Вадецкая Э.Б. Археологические памятники в степях Среднего Енисея. //Л., «Наука», 1986. 180 с. http://kronk.narod.ru/library/vadezkaya-1986.htm Азбелев П. Сайт-монография. Древние кыргызы. Очерки истории и археологии. http://kronk.narod.ru/
Деренко М.В., Малярчук Б.А. Генетическая история коренного населения Северной Азии. Природа. №10. 2002.
http://srv1.rsl.ru/VV/JOURNAL/NATURE/10_02/ASIAN.HTM
[Леонид Драчевский] Леонид Драчевский против возврата "скифской принцессы" алтайцам. http://regnum.ru/allnews/251678.html Сайт http://regnum.ru/ «Алтайская принцесса» оказалась скифской. http://www.midinfo.ru/cgi-bin/eng.pl?c=news&a=view&cat=syb&nid=509 Междуреченский информационный портал.
Молодин В., Ромащенко А. Палеогенетический анализ генофонда древнего населения Сибири. //Наука в Сибири №49. 1999. http://www-sbras.nsc.ru/HBC/1999/n49/f4.html Сайт газеты «Наука в Сибири» http://www-sbras.nsc.ru/HBC/about.php?lang=ru
Захаров И.А. Центральноазиатское происхождение предков первых американцев. Первые американцы. 2003. № 11. С. 139-144.
http://macroevolution.narod.ru/zaharov_indians.htm Сайт Проблемы эволюции.
http://macroevolution.narod.ru/index.html
[ХРОН4] Носовский Г. В., Фоменко А.Т. Новая хронология Руси. Русь. Англия. Византия. Рим. (В трех томах). Изд. РИМИС. 2005. //chronologia.org/xpon4/index.html Сайт проекта «Новая Хронология». //chronologia.org
[ХРОН5] Носовский Г. В., Фоменко А.Т. Империя. Изд. РИМИС. 2005. //chronologia.org/xpon5/index.html Сайт проекта «Новая Хронология». //chronologia.org
[ХРОН6] Носовский Г.В., Фоменко А.Т. Библейская Русь (в четырех томах). Изд. РИМИС. 2004 г. //chronologia.org/xpon6/index.html Сайт проекта «Новая Хронология». //chronologia.org
Полосьмах Н.В. Тайны «царских» Пазырыкских курганов. http://www.inauka.ru/analysis/article65347 Сайт «Известия науки» http://www.inauka.ru/
Райс Т.Т. Скифы: строители степных пирамид. 2004. Центрполиграф.
http://bibliotekar.ru/skify/index.htm
Сайт Новая Хронология. //chronologia.org/
[Сибирский энеолит] Сибирский энеолит и эпоха бронзы. http://sibirfilm.narod.ru/RUS/proekt-bronz.htm Проект «Тайная Сибирь». http://sibirfilm.narod.ru/RUS/proekt-rus.htm
Тихонов А.Г. Новые данные к антропологии населения кобанской культуры (по материалам могильника Уллубаганалы).
http://s155239215.onlinehome.us/turkic/62_Scythians_7BC/TikhonovUllubaganalyRu.htm
Сайт Turkwold. http://www.turkicworld.org/
[Тюрин, 2005, Алгоритмы] Тюрин А.М. Алгоритмы фальсификации и ре-фальсификации результатов радиоуглеродных датировок.
/volume3/turin_alg.html Электронный сборник статей «Новая Хронология». Выпуск 3. 2005. /volume3/ Сайт Новая Хронология. //chronologia.org/
[Тюрин, 2007, Интеграция, Скифы] Тюрин А.М. Интеграция скифской археологической культуры в Новую Хронологию Фоменко и Носовского. /volume6/tur_int.doc Электронный сборник статей «Новая Хронология». Выпуск 6. 2007. /volume6/index.html Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2007, Лингвистика, Скифы] Тюрин А.М. Казаки, черкасы, татары, орда и скифская археологическая культура. /volume6/tur_kaz.doc Электронный сборник статей «Новая Хронология». Выпуск 6. 2007.
/volume6/index.html Сайт: Новая Хронология.
//chronologia.org/
Тур С.С., Рыкунов М.П. Краниологические материалы пазырыкской культуры из могильников в урочище Кызыл-джар. Древности Алтая. №12. 2004. http://e-lib.gasu.ru/da/archive/2004/12/05.html Электронная библиотека ГАГУ. http://e-lib.gasu.ru/
Хуснутдинова Э. К. Этногеномика и генетическая история народов Восточной Европы. Вестник российской науки. том 73, № 7, с. 614-621 (2003).
http://vivovoco.rsl.ru/VV/JOURNAL/VRAN/03_07/ETHNOGENE.HTM
Ялбачева Н.М., Стрельцова Т.А. Генетический подход к происхождению коренных народов республики Алтай. http://altay-250-letie.ru/konfer/yalb_strel.html Электронное периодическое издание «Алтай.250-летие.ру». http://altay-250-letie.ru/news/news.html
Alekseev A. Yu., Bokovenko N. A., Boltrik Yu., Chugunov K. A., Cook G., Dergachev V. A., Possnert G., Plicht J., Scott E. M., Semeetsov A., Kovalyukh N., Skripkin V., Vasiliev S. and Zaitseva G. Chronology of Eurasian Scythian Antiquities Born by New Archaeological and 14C Data. Radiocarbon, Vol. 43, No 2B, 2001, p 1085-110. Proceedings of the 17th International 14C Conferenc.
http://s155239215.onlinehome.us/turkic/btn_Archeology/ScythianC14En.htm Сайт Turkic History. http://www.turkicworld.org/
Tur S.S. Modern Descendants of Pazyryk Culture Carriers. (Barnaul, 2003).
http://s155239215.onlinehome.us/turkic/64_Pazyryk/PazyrykTurCraniologyEn.htm Сайт Turkic History. http://www.turkicworld.org/ http://e-lib.gasu.ru/da/archive/2003/10/14.html
Voevoda M.I., Sitnikova V.V., Romashchenko A.G., Chikiseva T.A., Polos’mak N.V., Molodin V.I. Reconstruction of The Genofond Peculiarities of The Ancient Pazyryk Population (I-II Millennium BC) from Gorny Altai According to The MTDNA Structure.
http://www.bionet.nsc.ru/bgrs/thesis/99/index.html
http://s155239215.onlinehome.us/turkic/64_Pazyryk/Pazyryk_gensRu.html Сайт Turkic History. http://www.turkicworld.org/
статья получена в 2007г.
