Русь-Орда и генетические карты Старого Света
Тюрин Анатолий Матвеевич, к.г.-м.н.
В соответствии с Новой Хронологией А.Т. Фоменко и Г.В. Носовского, так называемая монгольская империя – это Русь-Орда, возникшая в 14 веке в Восточной Европе и начавшая отсюда экспансию на восток, юг, и запад. Военным инструментом экспансии являлась Орда – армия, сформированная и организованная на основе определенных принципов. Записаны ли этапы формирования и развития Руси-орды в генетических картах Старого Света? Да. Это общий вывод, который сделан на основе анализа генетических карт.Частные выводы сводится к следующему. По генетическим картам Старого Света, с привлечением другой генетической информации, возможно построение моделей-реконструкций, которые иллюстрируют гипотезы Новой Хронологии:
- в Руси-орде была создана специфическая военная структура – Орда;
- прямое влияние Руси-орды распространялось на обширные территории Европы, Азии и Северной Африки;
- Русь-Орда имела регулярные сообщения с Америкой;
- Византия и ее генетическая наследница Турция были единственными империями в Средиземноморье, просуществовавшими длительное время;
- Римская империя с центром на Апеннинском полуострове - фантом Традиционной Истории.
Три массива генетической информации – карты распределения групп крови, данные, обосновывающие наличие общего предка у славян и популяций Северного Индостана, а также характерное распределение европеоидности в кастах популяций Южного Индостана, прямо и однозначно голосуют за соответствие действительности одной из самых ярких гипотез НХ ФиН - арии пришли в Индию из Руси-Орды.
Ключевые слова: Новая Хронология, Русь-Орда, генетические карты, реконструкция.
Оглавление
1. Постановка задачи
2. Общие сведения
3. Карты главных компонент генетических кодовнародов Европы и Ближнего Востока
3.2. Первый компонент
3.3. Второй компонент
3.4. Третий компонент
3.5. Четвертый компонент
3.6. Пятый компонент
3.7. Отсутствующие компоненты
4. Элементы НХ-реконструкций прошлого Человечества по генетической информации
4.2. Русь-Орда и Европа
4.3. Русь-Орда и Америка
4.4. Русь-Орда и Азия
4.5. Русь-Орда и Египет
4.5. Русь-Орда и Индостан
4.6. Скандинавия
5. Состояние генетических реконструкций прошлого народов и социальных групп
6. Заключение
Источники информации
1. Постановка задачи
В соответствии с Новой Хронологией А.Т. Фоменко и Г.В. Носовского (НХ ФиН) [Сайт НХ ФиН], так называемая монгольская империя – это Русь-Орда, возникшая в 14 веке в Восточной Европе и начавшая отсюда экспансию на восток, юг, и запад. Военным инструментом экспансии являлась Орда – армия, сформированная и организованная на основе определенных принципов [ХРОН4, Глава 3]. Записаны ли этапы формирования и развития Руси-орды в генетических картах Старого Света? Поиску ответа на этот вопрос и посвящена настоящая статья.
2. Общие сведения
Различают две группы генетических маркеров человека. Генетические маркеры (гены), которые:
- не влияют на биологические особенности людей, то есть, не определяют их «конкурентоспособность» в выживании в определенных природных и социальных условиях;
- определяют биологические особенности людей и являются объектом естественного отбора.
К последним относятся и гены, которые ответственны за группу крови человека и ее резус-фактор.
В генетике имеется такое понятие, как эмиссия генов. Она может происходить, в основном, двумя путями. Первый – брачный обмен, второй – переселение части популяции на новое место жительства. Вот и всё, что нам понадобится для поиска ответа на вопрос, сформулированный в разделе «Постановка задачи».
3. Карты главных компонент генетических кодовнародов Европы и Ближнего Востока
3.1. Общие характеристики карт
В публикации [Cavalli-Sforza, 1997] приведены результаты специальной обработки генетических кодов коренных народов Европы и Ближнего Востока. Анализировались генетические вариации 95 классических полиморфных групп. Выделено 5 главных компонент (рисунок 1), обусловивших особенности их распределения в регионе, и даны версии их идентификации. Особенности конфигурации зон разных значтмостей главных компонентов объяснены на основе такого понятия как эмиссия генов. Ниже приведен критический анализ версий идентификации компонент автора публикации. Приведены так же наши версии их идентификации. При этом, мы придерживались простого правила: при идентификации главных компонент возможные факторы рассматривались в хронологическом порядке - от наших дней в прошлое. Это понятно. При прочих равных условиях наиболее контрастно в главных компонентах генетических кодовнародов Европы и Ближнего Востока проявились те факторы, которые действовали относительно недавно.
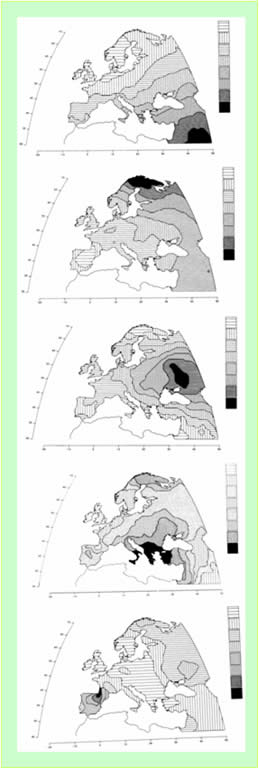
Рисунок 1. Карты главных компонентгенетических кодов коренных народов Европы и Ближнего Востока [Cavalli-Sforza, 1997]. Номера компонент – сверху вниз. Значимости компонент в обеспечении генетических вариаций: 28%, 22%, 11%, 7% и 5%.
3.2. Первый компонент
Первый компонент, обеспечивающий 28% генетических вариаций, по мнению автора публикации [Cavalli-Sforza, 1997], коррелируется с реконструированными по археологическим данным этапами распространения в регионе агрикультур в период 10000-6000 лет назад. В соответствии с этой версией, производящее сельское хозяйство возникло на Среднем Востоке и затем распространялось в северо-западном направлении. Вместе с его распространением мигрировали и гены людей, населявших Средний Восток. На наш взгляд, у этой версии идентификации Первого компонента имеется три «слабых» места.
1. Простирание изолиний значимости компонента и границ природно-климатических зон в Центральной и Северной Европе не соответствуют друг другу. В качестве примера можно привести схему зон сравнительной благоприятности для плодоводства (рисунке 2). В соответствии с версией автора публикации [Cavalli-Sforza, 1997] получается, что производящее сельское хозяйство распространялось по территории региона с постоянной скоростью, вне зависимости от простирания и конфигурации зон сравнительной благоприятности для плодоводства.
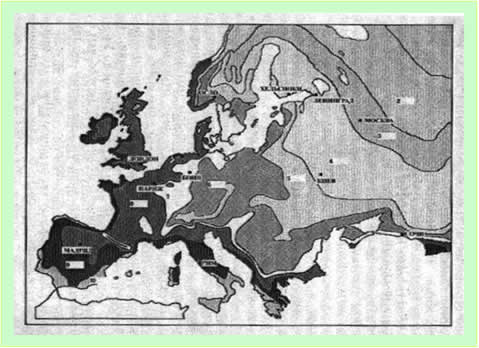
Рисунок 2. Схема зон сравнительной благоприятности для плодоводства (из книги Х. Бейкера «Плодовые культуры», М, Мир, 1990, дополнена по материалам книги Е. Ярославцева и др. «Ваш сад», М., Агропромиздат, 1992) [Паршев].
2. По статистической выборке данных радиоуглеродного датирования, включающей кости человека и материалы, характеризующих продукты его питания, нами построена модель «Европейцы голоцена» [Тюрин, 2005, Европейцы голоцена]. Особенностью выборки является включение в нее, наряду с радиоуглеродным возрастом образцов, и параметра D13С, отражающего относительное содержание в них стабильных изотопов углерода. Этот параметр интегрально характеризует диету человека. Возможно, люди в Европе появились не ранее примерно 12540 до года н.э.. Питались они либо как животные, либо только животными. Скорее всего, это было сообщество охотников. Примерно с 8200 года до н.э. в Европе начал сформироваться конгломерат сообществ охотников и собирателей. Для него характерны региональная унификация диеты при больших ее различиях между регионами. Это можно интерпретировать как региональное разделение сообществ охотников и собирателей и специализацию их диеты. Для части сообществ собирателей характерна диета типа морской/речной. Около 3800 года до н.э. конгломерат сообществ охотников и собирателей Европы практически мгновенно перестал существовать. Без видимой связи с конгломератом сообществ охотников и собирателей возникло сообщество земледельцев. Основная его характеристика – относительная унификация диеты в пределах всей Европы при сохранении устойчивых во времени региональных различий. Другими характеристиками сообщества земледельцев являются периодические сокращения его численности. Рубеж около 3800 года до н.э. можно принять за начало в Европе технологической цивилизации. Рассмотренные нами данные позволяют сделать предположение о вынужденном переходе европейцев на этот путь развития. Модель «Европейцы голоцена», построенная по фактическим данным, исключает возможность идентификации Первого компонента, с культурной эмиссией, протекающей относительно длительное время. Если судить по типу диеты, то смена хозяйственного уклада населения Европы произошла практически мгновенно.
3. Автор публикации [Cavalli-Sforza, 1997] идентифицировал Первый компонент на основе археологических данных, характеризующих распространение агрикультур.«Thus, in Europe, the most important hidden pattern (the first PC) has an extremely high correlation with the history of the spread of agriculture from the Middle East in the period 10,000-6,000 years B.P.» Здесь и далее индексом ВР обозначается некалиброванные радиоуглеродные даты (ВР – before present, present = 1950 год). Но археологические данные по распространению агрикультур, на которые он ссылается, кардинально не соответствуют этапам развития неолита в рассматриваемом регионе. «Накопление радиоуглеродных дат уже сейчас позволяет ставить проблемы, имеющие большое историческое значение. К их числу можно отнести проблему неолитизации: процесс зарождения и распространения хозяйства производящего типа; параллелизацию развития форм хозяйства и этапов развития природной среды; проблемы временного соотношения археологических общностей на значительных пространствах.» [Долуханов, 1972]. Что такое неолитизация можно понять по картам Европы и сопредельных регионов Азии, на которых для разных периодов показаны археологические культуры и природно-климатические зоны. В период 10500-8000 ВР годов культуры неолита были только в Греции, Анатолии, Сирии и Палестине [Долуханов, 1972, таблица 1]. Синхронными им были культуры мезолита в других частях Европы. В период 8000-7000 ВР годов зона культур неолита расширилась за счет территорий Югославии и Южной Италии [Долуханов, 1972, таблица 2]. Возникли очаги неолита в Закавказье, Прикарпатье и на юге Франции. В период 5500-4000 ВР годов неолит распространился по всей Центральной и Западной Европе [Долуханов, 1972, таблица 6]. Но в Греции и Анатолии в этот период археологические культуры неолита не показаны, что является следствием их радиоуглеродного датирования по реконструированному нами алгоритму радиоуглеродного датирования археологических культур неолита Евразии. Археологические культуры этого периода передвинуты в прошлое на 1620 лет. Культуры неолита в северном Причерноморье появились только в период 4500-4000 ВР годов [Долуханов, 1972, таблица 8]. В этот же период культуры неолита появились в Индии и Пакистане [Долуханов, 1972, таблица 4]. Графические схемы, иллюстрирующие процесс неолитизации, имеют еще одну особенность [Долуханов, 1972, таблицы 1-8]. Наиболее древний неолит представлен археологическими культурами докерамического неолита. Эти культуры выделены только в Восточном Средиземноморье. При построении графических схем эти культуры обозначены, как неолит. Исходя из этого, нижняя граница собственно неолита, например, Англии (примерно 5200 ВР год) идентифицирована с нижней границей докерамического неолита, например, Анатолии (последняя дата 9520 ВР год).
Отметим, что «слабое» место идентификации Первого компонента, отраженное в пункте 3, обозначено формально. Технически алгоритм радиоуглеродного датирования археологических культур неолита Евразии реализован за счет радиоуглеродного датирования на основе двух эталонов – 13,56 dpm/g и 15,3 dpm/g [Тюрин, 2005, Алгоритмы]. Их применение дает разный возраст одного и того же датируемого образца. Разница составляет 1620 радиоуглеродных лет. Более древний возраст получают при датировании на основе эталона 15,3 dpm/g. Датирование археологических культур Греции и Югославии на основе эталона 15,3 dpm/g начато с 4080 ВР года, Анатолии - с 5000 ВР года, Сирии и Палестины – с 5380 ВР года. За счет этого возраст археологических культур неолита этих исторических регионов искусственно увеличен на 1620 лет. Имеются признаки того, что этим же способом увеличен возраст части культур неолита Месопотамии. Возможно, увеличен возраст и части культур неолита Венгрии и Румынии. Археологические культуры неолита других регионов Евразии датированы на основе эталона 13,56 dpm/g. То есть, фактическая основа модели «Неолитизация» сфальсифицирована. А раз так, то мы не в праве требовать соответствия конфигурации изолиний значимости Первого компонента тому, что не соответствует действительности. Это так. Однако, подробное рассмотрение этого вопроса выполнено с конкретной целью. Декларация автора публикации[Cavalli-Sforza, 1997] о соответствии конфигурации изолиний значимости Первого компонента археологическим данным не соответствует действительности. Конфигурация изолиний как раз кардинально не соответствует археологическим данным, принимаемым сегодня за вполне достоверные факты.
В рассматриваемом регионе имеется две зоны контрастного проявления Первого компонента. Первая – максимальное его проявление в районе Ближнего Востока, вторая – минимальное проявление - в регионе, охватывающем Англию, Ирландию, Бельгию, Голландию, Данию, северо-запад Германии, южную Норвегию, южную Швецию и южную Финляндию. Что случилось в этом регионе в ближайшем прошлом? Промышленная революция Нового времени. На первом ее этапе произошло скачкообразное увеличение производительности сельского хозяйства, достигнутое и искусственным путем. При этом, экстенсивные возможности его развития в регионе были резко ограничены. Вместе с тем возникли промышленные центры, нуждающиеся в рабочих. Все это привело к значительной локальной миграции населения. С другой стороны, в регионе Среднего востока, основная месса населения до сих пор проживает в тех местах, где проживали их предки. Таким образом, Первый компонент отражает степень генетической однородности населения на локальном уровне. На Среднем Востоке она максимальна, в регионе, где произошла промышленная революция – минимальна. Главный фактор, обусловивший наличие Первого компонента – социальные последствия промышленной революции и развивающегося после нее капиталистического уклада хозяйства. Из региона, победившей промышленной революции началась ее экспансия. Ее главный вектор был направлен на юго-восток. Экспансия имела две принципиально разные составляющие: экспорт технологий, включающий и технологии организации производящего хозяйства, и экспорт специалистов (предпринимателей и рабочих). В первой половине 20 века промышленная революция достигла Ближнего Востока. В 21 веке она достигнет Среднего Востока. Этапы продвижения промышленной революции в юго-восточном направлении как раз и записаны в конфигурации изолиний значимости Первого компонента. Рассматриваемый процесс должен был иметь и другие социальные последствия. Он и имел. Область минимальных значений Первого компонента примерно соответствует распространению Христианства Протестантского толка. Область максимальных значений – Ортодоксального Ислама. Контрастность проявления Первого компонента обратно пропорциональна степени разнообразия полиморфизма. Это и создает иллюзию того, что Первый компонент связан с эмиссией генов со Среднего Востока в северо-восточном направлении. Хронологический интервал проявления Первого компонента – 17 век – современность.
3.3. Второй компонент
Второй компонент обеспечивает 22% генетических вариаций. Автор публикации [Cavalli-Sforza, 1997] предположительно идентифицировал его с миграцией на север Европы народов, говорящих на языках уральской семьи: лопарей, финнов, венгров и других народов. В российской литературе их называют угорскими народами. Отметим, что даже в специальной литературе наблюдается смешение двух понятий – типологизация языков, на которых говорят народы Мира, и их генетических особенностей. Безусловно, эти две характеристики коррелируются на глобальном уровне. Но на региональном уровне могут быть существенные отклонения от общих схем соответствия. Тюрки - наиболее яркий пример таких отклонений. Тюрки - это генетически далекие друг от друга нарды, но говорящие на языках, объединенных в одну семью. Поэтому мы не можем принять вышеобозначенную идентификацию Второго компонента.
Наиболее значимое проявление Второго компонента наблюдается в регионе к северу от Балтийского моря. Простирание изолиний значимости компонента и границ природно-климатических зон в Центральной и Северной Европе в целом соответствуют друг другу. По нашему мнению, фактором, с которым связан Второй компонент, является отступление ледников в начале голоцена. Наша версия сводится к следующему. В пре-голоценовое время сложилась общность людей, приспособившихся к жизни вблизи ледников. Естественно, возникли и характерные для них полиморфные группы генов. С началом отступления ледников в голоцене за ними мигрировала часть людей этой общности. Другая часть людей оставалась на прежних местах своего проживания и была ассимилирована соседями, пришедшими из более южной природно-климатической зоны. Так продолжалось до тех пор, пока люди, двигавшиеся за отступавшим ледником, не дошли до берегов северных морей. Поэтому наиболее контрастное проявление Второго компонента как раз и приурочено к узкой прибрежной полосе севера Европы. Конфигурация значимостей Второго компонента имеет две характерные особенности. В районах вблизи северо-западных и восточных склонов Альп имеются локальные максимумы. Они связаны, скорее всего, с общностями людей, приспособившихся к жизни вблизи кромки ледника, которые попали в естественную природную ловушку. Главный ледник отступил на северо-восток, а они остались жить около ледника, сформированного на Альпах. При потеплении климата этот ледник отступил в центральные районы Альп, а люди, проживавшие около его кромки, остались на приальпийских равнинах. Там они и были ассимилированы пришельцами. Таким образом, Второй компонент связан, главным образом, не с эмиссией генов, но с их смешением. Хронологический интервал проявления Второго компонента, как смешение – начало голоцена. В последствии Второй компонент проявлялся через эмиссию генов. Можно предположить, что относительно высокие значения Второго компонента на Ближнем и Среднем Востоке, связаны с эмиссией генов из Западной Сибири и Южного Урала.
3.4. Третий компонент
Третий компонент, по мнению автора публикации [Cavalli-Sforza, 1997] связан с возникновением кочевого хозяйства и распространением популяций, живущих за его счет. Компонент обеспечивает 11% генетических вариаций и имеет чрезвычайно выраженную форму изолиний значимости. Наиболее контрастное его проявление приурочено к области, охватывающей донские степи и территории, примыкающие к ним с севера. Москва находится или внутри зоны наибольшей значимости компонента или рядом с ее границей. Убывание значимости компонента происходит концентрически. Идентификация компонента с кочевниками, не выдерживает никакой критики. Кочевники могли сформироваться из того субстрата, который уже находился в Восточной Европе. А он к моменту возникновения кочевого хозяйства генетически был относительно однороден в пределах локальных областей всей Европы. Кроме того, конфигурация изолиний значимости компонента не соответствует границам природно-климатических зон. Область его наибольших значимостей включает участки зон степи, лесостепи и смешанного леса. Это говорит об искусственном формировании компонента. Третий компонент – это Орда Руси-орды. Более детально Третий компонент рассмотрен в разделе «Русь-Орда и Европа».
3.5. Четвертый компонент
Четвертый компонент (обеспечивает 7% генетических вариаций) проявился контрастно. Автор публикации [Cavalli-Sforza, 1997] однозначно идентифицировал его с греческой колонизацией 1 тысячелетия до н.э. По нашему мнению, такая идентификация не является ни правильной, ни не правильной. Она некорректна. Получилось так, что греческая колонизация 1 тысячелетия до н.э., как фактор, определяет генетический портрет сегодняшних европейцев и азиатов, а Византия, просуществовавшая в соответствии с Традиционной Историей (ТИ) сотни лет, никак на нем не отразилась. Наша идентификация Четвертого компонента тоже однозначна - это Византия. Более детально Четвертый компонент рассмотрен в разделе «Византия».
3.6. Пятый компонент
Пятый компонент (обеспечивает 5% генетических вариаций) идентифицирован автором публикации [Cavalli-Sforza, 1997] с общностями, говорящими на баскском языке. Мы это тоже не можем принять, поскольку значимости компонента имеют четыре локальных максимума. Самый контрастный из них действительно корреспондируется с зоной проживания басков и распространения их языка (рисунок 5). Три других локальных максимума значимостей компонента не имеют к баскам никакого отношения. Самый южный максимум «накрывает» исторические области Сирии и Палестины. Самый северный – крайнюю северную часть Европы. Еще один максимум практически совпадет с первичной областью формирования в 14 веке Орды Руси-орды (рисунок 3). По нашему мнению, общим знаменателем четырех локальных максимумов Пятого компонента является Америка. То есть, локальные максимумы значимостей Пятого компонента – это области, через которые гены коренных американцев в течении определенного времени попадали в Европу. Более детально Пятый компонент рассмотрен в разделе «Русь-Орда и Америка».
3.7. Отсутствующие компоненты
В главных компонентахгенетических кодов коренных народов Европы и Ближнего Востока не проявилось два значимых (в соответствии с ТИ) фактора: античная Римская империя с центром на Апеннинском полуострове и средневековая Римская империя с центром в Вене.
4. Элементы НХ-реконструкций прошлого Человечества по генетической информации
4.1. Византия
В соответствии с НХ ФиН Византийская империя возникла в 10-11 веках. В 13 веке она вступила в период упадка. В 14 веке имперский центр Византии оказался под контролем Руси-орды, а в 15 веке он превратился в цент другой империи – Атаманской, созданной при непосредственном участии Руси-орды. Атаманская (турецкая) империя просуществовала до начала 20 века. Таким образом, в НХ ФиН четко различаются два понятия: Византийская империя и ее имперский центр [Носовский, 1995, Империя]. Возникнув в 10-11 веках, имперский центр просуществовал практически до 19 века включительно, то есть примерно 1000 лет. Никаких серьезных потрясений, которые могли исказить генетический портрет имперского центра Византии-Турции, в 20 веке не произошло. Значит, имперский центр Византии просто обязан проявиться вгенетических кодах коренных народов Европы и Ближнего Востока. Он и проявился. Имперский центр Византии (или ее генетическое ядро) проявился как максимальные значимости Четвертого компонента. Генетическое ядро Византии охватывает Балканы, западную часть Малой Азии, южную часть Апеннинского полуострова и Сицилию (рисунок 3). Генетическую границу Византии можно провести по изолинии, разделяющей третью и четвертую градации значимостей компонента. В генетические границы Византии попадают юг Франции, Италия, Центральная Европа, Северное Причерноморье и Ближний Восток. Имеется локальная область высоких значимостей Четвертого компонента – регион к северу от Балтийского моря. Что общего у народов, населявших этот регион и народов Византии? Этот вопрос мы оставим пока без ответа. Только отметим, что генетическая контрастность народов, проживающих к северу от Балтийского моря, требует специального рассмотрения. Генетические границы Византии на северо-востоке непосредственно соприкасаются с исконно русскими областями. Побережье восточной части Черного моря в генетические границы Византии не входит. В центральной части Пиренейского полуострова имеется локальный максимум значимости Четвертого компонента. Возможно, он связан с миграцией в эту область части населения Константинополя, после его захвата в 1452 году турками.
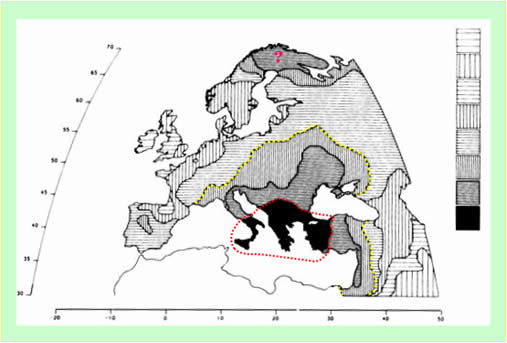
Рисунок 3. Четвертый компонентгенетических кодов коренных народов Европы и Ближнего Востока [Cavalli-Sforza, 1997].
Точечная линия красного цвета - границы генетического ядра Византии.
Точечная линия желтого цвета – генетические границы Византии.
4.2. Русь-Орда и Европа
На начальном этапе Орда формировалась за счет призыва в нее молодежи с территории Руси-орды [ХРОН4, Глава 3]. Со временем Орда превратилась в самопроизводящую сословную структуру.Естественно, самопроизводился и ее генетический портрет. Если это так, то главным антропологическим и генетическим маркером Орды будет, прежде всего, европеоидность, которая сложилась в Восточной Европе в середине 2 тысячелетия н.э. Вторым маркером Орды будет высокий уровень антропологического и генетического разнообразия входящего в нее военного сословия. Именно это генетическое разнообразие проявилось как Третий компонентгенетических кодов коренных народов Европы и Ближнего Востока. Это разнообразие качественно отличается от разнообразия, сформированного при смешении населения на локальном уровне и отраженного Первым компонентом. Генетическое разнообразие Орды сформировано на региональном уровне. Естественно, что минимальные значения значимостей компонента приурочены к областям, где не имелось условий для формирования генетического разнообразия на региональном уровне. К ним относятся южные части Балканского и Апеннинского полуостровов, Сицилия, Пиренейский полуостров и регион, расположенный к северу от Балтийского моря. В соответствии с ТИ на территории Палестины и Иордании происходили бурные исторические события, следовательно, проживающие на ней популяции должны иметь высокую степень регионального генетического разнообразия. Фактически же популяции этой территории характеризуются минимальными значениями этого параметра. Это и следует принять за основу при выполнении НХ-реконструкций истории Человечества.
Генетический портрет Орды распространялся тремя способами:
- расширение области базирования подразделений Орды (14-16 века);
- расположение подразделений Орды на завоеванных Русью-ордой территориях (14-16 века)
- эмиссия генов (14 век – современность).
Результаты нашей «количественной» географической реконструкции «Русь-Орда и Европа», выполненной на основе идентификации Третьего компонента генетических кодов, сводятся к следующему. В 14 веке Орда была сформирована в зоне, включающей донские степи. Это одно из самых удобных мест для постоянного расположения регулярной армии Руси-орды: обилие и высокое качество травянистых растений для корма животных, мягкий климат, равноудаленность от земель, которые планировалось завоевать в первую очередь. Одна из ставок правителей Руси-орды – Семи-Каракорум, находилась в низовьях Дона. Строительство в 15 веке новой столицы Руси-орды – Москвы, решало задачу ее (столицы) географического приближения к Орде. Москва и ее окрестности были заселены в 15-16 веках, в основном, генетическими ордынцами. Это и обусловило характерную конфигурацию области максимальных значимостей Третьего компонента. Она имеет как бы вытянутый выступ накрывающий (или почти накрывающий) Москву (рисунок 4). Примерно, на рубеже 14 и 15 веков границы области расположения Орды были расширены за счет включения в нее Северного Причерноморья и Северного Прикаспия. Генетические границы Руси-орды можно провести по изолинии, разграничивающей пятый и шестой уровни значимости Третьего компонента. На западе в них попадают юг Норвегии, юг Швеции, Дания и Германия. То есть получилось, что Балтийское море являлось внутренним морем Руси-орды. Генетические границы Руси-орды охватывают области распространения славянских и германских (за исключением Англии и Ирландии) языков (рисунок 5).
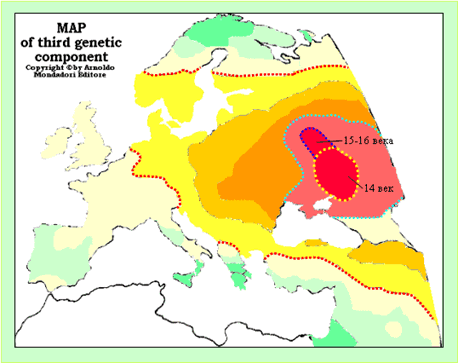
Рисунок 4. Третий компонентгенетических кодов коренных народов Европы и Ближнего Востока [Cavalli-Sforza, 1997]. Ярко красный цвет – область наибольшей значимости компонента, бирюзовый цвет – наименьшей значимости. Точечная линия желтого цвета - границы первичной области формирования в 14 веке Орды Руси-орды. Точечная линия синего цвета – граница области (Москва и Подмосковье), заселенной в 15-16 веках генетическими ордынцами. Точечная линия голубого цвета – граница области расположения Орды в 15-16 веках. Точечная линия красного цвета – генетические границы Руси-орды.
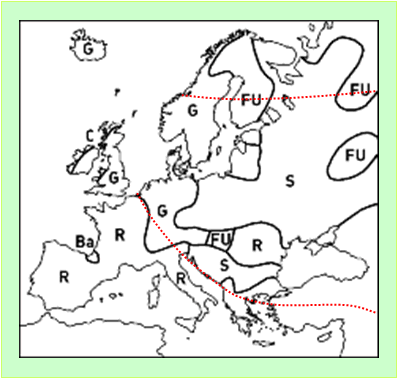
Рисунок 5. Схема распространение языков в Европе в настоящее время. (Ba = баскские, C = кельтские, FU = финно-угорские, G = германские, R = романские, S = славянские языки) [Древнейший язык].
Точечная линия красного цвета – генетические границы Руси-орды.
4.3. Русь-Орда и Америка
В НХ ФиН четко обозначена связь древних цивилизаций Америки и Руси-орды. «В архитектуре мексиканских «древних» пирамид и дворцов тоже отчетливо прослеживаются связи с европейскими русско-ордынскими пирамидами-курганами.» [ХРОН5, часть 5]. Высказана гипотеза о том, что Мексика являлась одной из земель Руси-орды. Так ли это? Попытаться найти ответ на этот вопрос можно на основе предельно простой логической конструкции. Считается, что из Европы совершали регулярные плавания в Америку, начиная с конца 15 века. Кроме всего прочего из Америки в Европу привозили гены американских индейцев и их болезни. Болезни действовали на людей с разными генами по-разному, производя специфический естественный отбор. То есть, в Европе, в регионах, из которых регулярно плавали в Америку, у современных людей должны иметься специфические генетические маркеры. Можно считать фактом, что одним из таких регионов является страна басков. Значит, у современных басков должны быть специфические американские генетические маркеры и их можно увидеть на соответствующих генетических картах. На этих же картах можно поискать регионы, где у современного населения встречаются такие же маркеры. Если, по крайней мере, один из таких регионов окажется на территории Руси-орды, то это будет означать, что гипотеза об американских владениях Руси-орды подтверждена фактическими генетическими данными. Результаты интерпретации генетических карт - схемы-реконструкции маршрутов регулярных плаваний из Европы и с Ближнего Востока в Америку, показаны на рисунке 6.Одним американским генетическим маркеромявляется максимальные значимости Пятого компонента генетических кодов. Наша реконструкция, выполненная на основе распределения значимостей этого параметра, показаны на рисунке 6, А.Еще одним американским генетическим маркеромявляются минимальные значения частот отрицательного резус-фактора крови. Мы предполагаем, что общая картина распределения частот отрицательного резус-фактора в Европе и на Ближнем Востокеобусловлена естественным отбором. Фактор естественного отбора – специфические американские болезни, летальный исход от которых у людей с отрицательным резус-фактором был выше, чем у людей с положительным. Из плаваний в Америку возвращался меньший процент людей с отрицательным резус-фактором, чем с положительным. Кроме того, американские болезни распространялись и в центрах, из которых осуществлялись регулярные плавания в Америку. Это тоже меняло процентное соотношение проживающих в них людей с отрицательным и положительным резус-факторами крови. На основе этих предположений по частотам отрицательного резус-фактора выполнена еще одна реконструкция маршрутов регулярных плаваний из Европы и с Ближнего Востока в Америку (рисунке 6, Б).
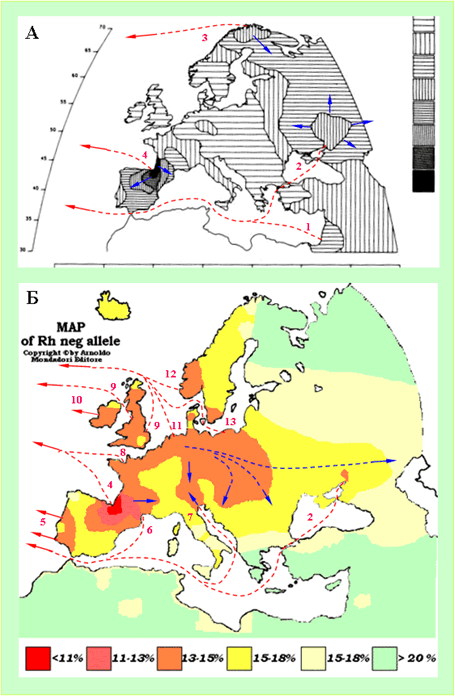
Рисунок 6. Схемы-реконструкции маршрутов регулярных плаваний из Европы и с Ближнего Востока в Америку. А - Пятый компонентгенетических кодов коренных народов Европы и Ближнего Востока [Cavalli-Sforza, 1997]. Б – схема частот отрицательного резус-фактора крови у популяций Европы, Ближнего Востока и Северной Африки [http://s155239215.onlinehome.us/turkic/63_Blood_Types/Blood%20TypesRu.htm#Группы%20Крови%20В]. Пунктирными линиями и стрелками красного цвет показаны маршруты регулярных плаваний в Америку. Пунктирными линиями и стрелками синего цвета - главные направления эмиссии генов. А – первая половина и середина 2 тысячелетия н.э.. Б – середина и вторая половина 2 тысячелетия н.э.. Красными цифрами пронумерованы центры, из которых осуществлялись регулярные плавания в Америку: 1 -Сирия и Палестина; 2 – низовья Дона; 3 – северная Норвегия; 4 – страна басков; 5 – Португалия; 6 – Южная Франция; 7 – северная Адриатика; 8 – Бретань; 9 – Англия; 10 – Ирландия; 11 – Бельгия, Голландия, северная Германия; 12 – южная Норвегия; 13 – западная Балтика.
Центры, из которых осуществлялись регулярные плавания в Америку, выделены по двум независимым генетическим параметрам:«Пятый компонент» и «Резус-фактор». Центр вСирии и Палестине, скорее всего, был первым, но просуществовал он недолго. Об этом говорит его выраженность только по параметру «Пятый компонент». Центррегулярных плаваний в Америку из Руси-орды находился в низовьях Дона. Этот центр контрастно проявился по параметру«Пятый компонент». Из него американские гены распространились по всей первичной области формирования в 14 веке Орды (рисунок 6), а так же за ее пределы. Вполне значимо центр проявился и по параметру «Резус-фактор». Возможно, этот центр регулярных плаваний в Америку включал и Крым.Однако, в 18 веке христианское население Крыма, являющееся в основном носителями американских генетических маркеров, было из него выведено и поселено опять же в низовьях Дона, вернее, в северном Приазовье. Строительство ПетромI флота в Воронеже, скорее всего, осуществлялось не на пустом месте. Верховья Дона с его сосновыми лесами являлось в 14-16 веках одной из баз строительства флота Руси-орды, в том числе и трансокеанского. А главной целью неудачного Азовского похода ПетраI являлось восстановление контроля над географическим регионом, в котором можно было бы воссоздать одну из самых мощных морских баз Руси-орды.
Центр регулярных плаваний в Америку, расположенный в Северной Скандинавии проявился только по параметру«Пятый компонент». До резкого похолодания в начале Малого Ледникового Периода - 1560 год [Тюрин, 2007, Датирование, МЛП], это было вполне приемлемым местом (географически, климатически, «ресурсно») для существования мощного морского промыслового центра. Из него же осуществлялись регулярные плавания в Америку. После 1560 года климатические условия в регионе резко ухудшились, что и привело к деградации промыслового центра. Можно ли считать, что центр сообщений с Америкой в Северной Скандинавии прямо подчинялся Руси-орде? Скорее да, чем нет. В любом случае этот промысловый центр мог существовать только «внутри» развитой торговой инфраструктуры, которая была создана в Европе в 14 веке Русью-ордой.
Наиболее интенсивное сообщение с Америкой осуществлялось из страны басков. Этот центр проявился как самые большие значения параметров «Пятый компонент» и «Резус-фактор». Другие европейские центры регулярных плаваний в Америку проявились только по параметру«Резус-фактор». Это связано, скорее всего, с тем, что народы, на территории которых они расположены, не имели привычки жениться на туземках, значит их потомки, с примесью американских генов, не попадали в Европу.
4.4. Русь-Орда и Азия
Орда, военная составляющая Руси-орды, была сформирована в 14 веке в бассейне Дона. В 14-15 векахобласть базирования подразделений Орды была расширена. Общие границы региона ее базирования реконструированы нами при интеграции скифской археологической культуры в НХ ФиН [Тюрин, 2007, Интеграция, Скифы]: степная и лесостепная зоны Евразии от северного Причерноморья до Саяно-Алтайского региона. В этом регионе формировались подразделения Орды, которые впоследствии были оставленына завоеванных Русью-ордой территориях. Из этого региона происходила эмиссия генов-маркеров Орды. Но был еще один фактор, обусловивший миграцию генетических маркеров Орды на восток, прежде всего, в Китай. Это кризис Орды в 17 веке, обусловленный ее тяжелыми военными поражениями в династических войнах за московский престол (в соответствии с ТИ: смута начала 17 века, и крестьянская война 1667-1671 годов). Одним из его специфических проявлений был «исход» вождей и людей военного сословия бывших орд Руси-орды в Китай, с образование там государства ордынского типа. «После захвата власти на Руси Романовыми в результате кровопролитной гражданской войны и разгрома Русской Великой Орды, оставшиеся в живых представители прежней Ордынской династии бежали в разные стороны. В частности, и на ВОСТОК. Некоторые, впрочем, пытались вернуться на московский престол. По-видимому, именно к таким попыткам следует отнести известное «восстание» Степана Разина, а потом «восстание» Емельяна Пугачева. Но кто-то бежал на Восток. Самой восточной Ордой была Пегая Орда, расположенная в то время вдоль границ современного Китая. Возможно, земли, занятые этой Ордой, и назывались КИТАЕМ. А современный Китай - ЧИНОЙ, как мы видели у Афанасия Никитина. Сбежавшая группа ордынцев была немногочисленной. Это и были манжуры = «монголы». Видимо, они везли с собой МАЛОЛЕТНЕГО ЦАРЕВИЧА. Кстати, в восстании Разина тоже фигурировал некий загадочный для историков ЦАРЕВИЧ Алексей. Набрав войско в Пегой Орде, они захватили Китай, обосновались там и предприняли все меры, чтобы не быть поглощенными романовской Россией. Для этой цели и была, в частности, создана Китайская Стена в середине или в конце XVII века. В 1644 году, как нам рассказывает скалигеровская история, они захватили, а скорее всего ОСНОВАЛИ, город Пекин. Или, как его тогда называли ПЕЖИН, от слова ПЕГАЯ Орда. Они провозгласили императором малолетнего царевича ШИ, которого привезли с собой из ЗОЛОТОЙ ОРДЫ, то есть с ВОЛГИ. Где в то время воевал Степан Разин. Войско завоевателей ушло в поход почти без женщин. Поэтому сохранять чисто манжурское = «монгольское» происхождение мог разве только императорский двор.» [ХРОН5, часть 2, глава 6].
Таким образом, в реконструкциях НХ ФиН имеется предположение, что в 1671 году «Войско завоевателей ушло в поход почти без женщин.» По результатам выполненного нами анализа данных, приведенных в многочисленных публикациях, мы сделали другое предположение[Тюрин, 2007, Интеграция, Скифы]: «исход» военного сословия пост-Ордынских формирований в Китай в 17-18 веках носил, скорее всего, массовый характер. То есть, в Китай ушли руководители и воины пост-Ордынских формирований, вместе со своими семьями и скотом. Об этом говорит, например, то, что Прикаспийские степи в начале 17 века опустели. Поэтому, в первой трети 17 века туда смогли откочевать калмыки из западной Монголии и Синьцзяна с образованием в регионе Калмыцкого ханства [Екеев, 2003]. В публикациях отмечается массовый «вывод» в 18 веке населения с Алтая [Маточкин, 1999] и Тувы [Боронин]. Население «выводилось» со скотом, а из Тувы в начале 18 века енисейские кыргызы «выведены» и с частью зависимых от них социальных групп. В середине – второй половине 18 века из праалтайских степей в Джунгарию ушли ак-теленгиты вместе с зависимыми от них социальными группами [Этнический состав]. Скорее всего, это был добровольный уход воинов на службу в маньчжуро-китайскую армию. Здесь важно то, что на службу в Китай уходили не просто воины, а вся государственная структура. «Трагическим стал 1703 год, когда джунгары угнали с родины около трех тысяч хакасов. Среди захваченных оказалось особенно много князей и правителей из числа царствующего рода Хыргыс, чиновников до того исправно действовавшего государственного аппарата. Началась агония Хакасского государства, правящая династия которого просуществовала с середины I в.до н.э. до 1703 г., т.е. около 1750 лет.» [Казласов, 2003]. Автор публикации [Нанзатов] упоминает потомков кыргызов, проживающих в Китае «Как отмечают Чжан Тайсянь, А.А.Шомаев и С.Г.Скобелев об этом свидетельствует большое количество архаизмов в языке фуюйских киргизов, проживающих в провинции Хэйлунцзян КНР и переселенных туда монголами.» Горнозаводчики отмечали в 20-х годах 18 века запустение Горного Алтая [Бородаев, 2003]. Таким образом, в 17-начале 18 века территории проживания, прежде всего военного сословия пост-Ордынских образований, от низовьев Волги до верховьев Енисея оказались заброшенными. Мы предполагаем, что именно этот фактор – уход военного сословия в Китай и обусловил то, что правительству Московского царства, а затем Российской империи пришлось снова «осваивать» Сибирь. Если все это так, то генетические маркеры Орды должны быть и в Китае.
Генетический вопрос «Русь-Орда и Азия» рассмотрен по картам частот групп крови В, А и О у популяций людей[DistributionofBlood]. Скорее всего генетическим маркером Орды являются пониженные частоты группы крови О (рисунок 7, В). Этот маркер Орды распространился в пределах огромной зоны, включающей Восточную Европу, Балканы, Ближний Восток, Египет, Среднюю Азию, Западную Сибирь, север Индостана, Тибет, Северный Китай и Приамуре. Фактически, зона пониженных частот группы крови О маркирует границу региона, завоеванного Русью-ордой в 14-15 веках, а также выходцами из нее в 17-18 веках. Высокие частоты группы крови В характерны, скорее всего, только для восточной части региона базирования Орды (рисунок 7, А). Граница высоких частот группы крови В в Азии практически совпадают с границей области пониженных частот группы крови О. В комментарии нуждается только один момент. Частоты этой группы на севере Индостана выше, чем в регионе базирования подразделений Орды. Группа крови – определяющий фактор естественного отбора людей. Исходя их этого, можно предположить, что на севере Индостана в период после его завоевания Русью-ордой более приспособленными для жизни в его климатических и социальных условиях оказались люди с группой крови В. Это же самое можно сказать для двух областей с наиболее высокими частотами группы крови В в Западной Сибири. Высокие частоты группы крови А характерны для Западной и Центральной Европы, Скандинавии, Балкан и современной территории Турции. Скорее всего, частоты группы крови А нейтральны по отношению к генетическому портрету Орды. Тем не менее, высокие частоты группы крови А были «транспортированы» Ордой из Европы в Азию, прежде всего в Северный Китай и Приамурье.
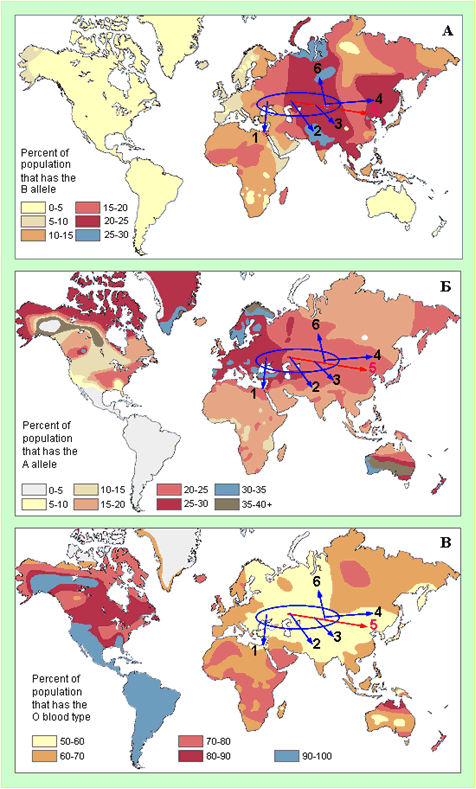
Рисунок 7. Карты частот групп крови В (секция А), А (секция Б) и О (секция В) у популяций людей[DistributionofBlood]. Голубой овал – регионбазирования подразделений Орды. Стрелки синего цвета – направления распространения генетического портрета Орды за счет расположение ее подразделений на завоеванных Русью-ордой территориях и эмиссии генов. Стрелки красного цвета - направления распространения генетического портрета Орды за счет «исхода» ордынцев в Китай.
4.5. Русь-Орда и Египет
Особенностью Руси-орды являлось то, что родовое кладбище ее правителей находилось в Египте [ХРОН5, часть 2, глава 19]. Египет завоеван Русью-ордой в 14 веке и в нем были сформированы войска, называемые мамелюками. «Мамелюки, как мы теперь понимаем, были отнюдь не «рабы, случайно захватившие власть», а остатки войск, охранявших царское кладбище и казну Империи в Египте. Надо думать, сюда посылались самые отборные воинские части Империи. После распада Империи в начале XVII века они оказались брошены на произвол судьбы. Тем не менее, еще долгое время они продолжали существовать самостоятельно и удерживать власть в Египте.» [Носовский, 2006]. В этой публикации приводятся и другие сведения, которые следует принять во внимание при реконструкции связей Руси-орды и Египта по генетическим данным. Мамелюки пополнялись в основном из подразделений Орды, расквартированных на Северном Кавказе. Владычество мамелюков в Египте завершилось в начале 19 века году, после того, как их армия была разбита Наполеоном. В 1811 году знатные мамелюки были истреблены. Если это так, то генетические маркеры Руси-орды должны присутствовать у народов, населяющих современный Египет и сопредельные страны. Карта частот группы крови В у популяций людей в Европе и сопредельных регионах (рисунок 8) прямо иллюстрирует одно из положений НХ ФиН: популяции Египта в 14-18 веках были тесно связаны с популяциями Дона и Северного Кавказа. Это же самое можно сказать и по карте частот группы крови О (рисунок 7). Следует отметить, что связь мамелюков с народами Северного Кавказа признается и в ТИ.
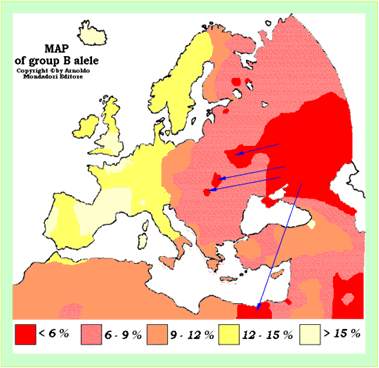
Рисунок 8. Карта частот группы крови В у популяций людей в Европе и сопредельных регионах [Сайт Turkic History].Внимание, условные обозначения, скорее всего, не соответствуют схеме. Красный цвет - > 15%. Синими стрелками показаны направления миграции популяций с высокими частотами крови группы В.
4.5. Русь-Орда и Индостан
«Многие исследователи обращали внимание на тесную связь между Индией и Русью. Теория о том, что арии, основавшие индийскую цивилизацию, пришли с севера, давно обсуждается в научной литературе. Наши исследования подтверждают данную точку зрения и показывают, что арии пришли в Индию из Руси-Орды в XIV-XV веках н.э., уже после Куликовской битвы.» [Носовский, 2007, Казаки-арии, Аннотация]. Карты частот групп крови подтверждают эту гипотезу. Имеются ли другие ее генетические подтверждения? Да. В публикации [Колёсов, 2007] отмечена близость генетических портретов славян и жителей Северного Индостана. «Так вот, и восточные славяне, и индусы (пакистанцы) принадлежат к одной и той же гаплогруппе – R1a. А значит, общий предок у них БЫЛ, ...». Скорее всего, гены «общего предка» занесены на Индостан подразделениями Орды, которые были оставлены в регионе после его завоевания Русью-ордой. В публикации [Колёсов, 2007] приводятся генетические данные по мусульманской части Индостана. Генетические данные, характеризующие индуисткий Индостан, приведены в публикации [Bamshad, 2001]. Генетические исследования выполнены по тщательно разработанной программе. При этом, все генетические данные были структурированы по индусским кастам (низшие, средние и высшие касты). Генетические портреты каст сопоставлены с генетическими портретами азиатов, европеоидов и африканцев. По полученным результатам сделаны следующие выводы. Доля европеоидности, особенно, европеоидности восточноевропейского типа, возрастает от нижних каст к высшим. («Moreover, the affinity to Europeans is proportionate to caste rank, the upper castes being most similar to Europeans, particularly East Europeans.»). При этом, европеоидность индусов по мужским линиям выше, чем по женским. Авторы публикации объяснили последнюю особенность тем, что мужчины составляли большинство эмигрантов (на Индостан) из западной Евразии (... suggested that the majority of immigrating West Eurasian may have been males.»). Самая высокая европеоидность у кшатриев, касты воинов. Также отмечено, что опубликованные гипотезы о миграции популяций из Европы на Индостан не имеют достоверных археологических подтверждений. Эти фактические данные однозначно подтверждают НХ ФиН в части завоевания в 14 веке Индостана войсками Руси-Орды. Ее воины сформировали высшие касты индусов, которые сохранились до наших дней.
Три массива генетической информации – карты распределения групп крови, данные, обосновывающие наличие общего предка у славян и популяций Северного Индостана, а также характерное распределение европеоидности в кастах популяций Южного Индостана, прямо и однозначно голосуют за соответствие действительности одной из самых ярких гипотез НХ ФиН - арии пришли в Индию из Руси-Орды.
4.6. Скандинавия
С началом генетических исследований выявился неожиданный факт. « ...Северная Европа испытала сильное генетическое влияние Восточной Сибири. Так, предками по мужской линии 52,4% финнов были монголоидные популяции Южной и Восточной Сибири. У эстонцев процент монголоидной мужской хромосомы не намного меньше. Значительный процент монголоидных отцовских линий того же происхождения, что и у финнов, согласно последним исследованиям имеется также у литовцев и латышей - и это несмотря на их индоевропейский язык. Несколько меньший - у норвежцев и шведов.» [Джадан, 2007]. Из этого факта сделан следующий вывод: «Это означает, что их праотцы около 5 тысяч лет назад жили в Забайкалье и на Алтае и были типичными монголоидами.» Наши реконструкции представлений о прошлом народов, выполненные в рамках НХ ФиН и приведенные в настоящей статье, а также в публикации [Тюрин, 2007, Интеграция, Скифы], позволяют обозначить гипотезы и о том, что причина возникновения рассматриваемого феномена связана с Русью-ордой. Одна из них – монголоидные гены в Скандинавию занесены из Америки. Новейшие данные о родственности генетических портретов коренных американцев и народов Алтая и Забайкалья приведены в статье [Тюрин, 2007, Биология, Скифы], а также в публикации [Карафет, 2006]. То есть, можно предположить, что гены из Алтая и Забайкалья попали в Америку в период 17-10 тысяч лет назад. А во втором тысячелетии н.э. эти гены попали в Скандинавию через расположенный на ее территории центр регулярных плаваний в Америку. Вторая гипотеза построена на аналогии. Если выходцы из Руси-орды смогли завоевать Китай и создать там государство ордынского типа, то они могли завоевать и север Скандинавии. Нами высказано предположение, что одной из основных задач Пегой орды, базировавшейся в Саяно-Алтайском регионе, был сбор ясака с народов Сибири [Тюрин, 2007, Интеграция, Скифы]. Скорее всего, существовали и спецподразделения, осуществлявшие сбор ясака в Западной Сибири. Возможно во время кризиса Орды (17 век) часть этих подразделений перебазировалась на север Скандинавии, и принесла сюда «алтайские» гены. Сегодня их прямыми потомками являются саамы. Обозначенные в наших гипотезах пути появления монголоидных генов в Скандинавии мы считает возможными с «качественных» позиций. Про то, как они согласуются с высоким процентом монголоидности финнов и эстонцев (с «количественными» характеристиками), мы ничего сказать не можем.
5. Состояние генетических реконструкций прошлого народов и социальных групп
Состояние генетических реконструкций имтории народов и социальных групп можно оценить по подборке материалов «ТИПЫ КРОВИ», опубликованной на сайте [Сайт Turkic History]. В одной из основополагающих монографий по этому вопросу в разделе «История человечества по генам» приведена следующая фраза: «Прежде всего, мы должны понять, что гены O, A, и B являются очень древними. Они не являются результатом недавних мутаций. Например, анализ египетских мумий показывает, что три гена, отвечающих за образование групп крови, присутствуют там в таких же пропорциях, как и у современных египтян ...» То есть генетики дают сами себе и своим читателям установку: основа интерпретации генетической информации – ТИ, в соответствии с которой египетские мумии относятся к 1-2 тысячелетиям до н.э. Исходя из этого, и следует формировать свои представления о скоростях генетических мутаций. В соответствии же с НХ ФиН египетские мумии следует датировать 14-18 веками н.э. [ХРОН5, часть 2, глава 19]. То есть, египетские мумии в своей массе просто обязаны быть генетически идентичными современным египтянам. Исключение составляют мумии фараонов, по крайней мере, часть которых принадлежит правителям Руси-орды, генетически не связанных с народами Египта. Скорее всего, ориентирование на ТИ при интерпретации генетических данных создало вгенетике целые направления «в никуда». Самое неприятное в этой ситуации – это то, что направления «в никуда» имеют обратную связь с массивом фактических генетических данных. На основе «в никуда» ведется структурирование массива, и планирование работ по его пополнению новыми данными. На основе «в никуда» создаются методики обработки и интерпретации генетической информации. Сегодня мы без труда нашли результаты «безыдейной» обработки фактических генетических данных - [Cavalli-Sforza, 1997]. Следует обратить внимание на то, что они были опубликованы 10 лет назад. Поэтому нам и удалось увидеть в них элементы, подтверждающие исторические реконструкции НХ ФиН. Но со временем, информационная база генетики в части реконструкций истории народов и социальных групп окажется «встроенной» в ТИ и настолько запутанной, что без специальных знаний в ней будет невозможно разобраться.
6. Заключение
По генетическим картам Старого Света, с привлечением другой генетической информации, возможно построение моделей-реконструкций, которые иллюстрируют гипотезы НХ ФиН:
- в Руси-орде была создана специфическая военная структура – Орда;
- прямое влияние Руси-орды распространялось на обширные территории Европы, Азии и Северной Африки;
- Русь-Орда имела регулярные сообщения с Америкой;
- Византия и ее генетическая наследница Турция были единственными империями в Средиземноморье, просуществовавши длительное время;
- Римская империя с центром на Апеннинском полуострове - фантом ТИ;
- арии пришли в Индию из Руси-Орды.
Общий вывод состоит из одного слова, являющегося ответом на вопрос, сформулированный в разделе «Постановка задачи». Записаны ли этапы формирования и развития Руси-орды в генетических картах Старого Света? Да.
Бородаев В.Б., Контев А.В. Взаимоотношение русских первопроходцев с монгольским и тюркском населением Верхнего Обь-Иртышья в период начального освоения рудных богатств Алтая (1720-е годы). http://new.hist.asu.ru/biblio/zalk/40-53.pdf Центральная Азия и Сибирь. Первые научные чтения памяти Е.М. Залкинда. 2003 г. http://new.hist.asu.ru/biblio/zalk/ Интернет-портал по истории Алтая. http://new.hist.asu.ru/
Боронин О.В. «Снятие» проблемы двоеданничества и двоеподданства
коренного населения бассейна Верхнего Енисея в первой половине XVIII в.
http://new.hist.asu.ru/biblio/borod3/403-407.html#text13 Интернет-портал по истории Алтая http://new.hist.asu.ru/
Джадан И. Похищение Евразии. Русский Журнал. 15 октября 2007 г. http://www.russ.ru/politics/docs/pohischenie_evrazii Сайт «Русский Журнал».
http://www.russ.ru/
[Древнейший язык] Древнейший язык в Европе.
http://www.terijoki.spb.ru/history/tpl3.php?page=wiik&lang=ru
Сайт http://www.terijoki.spb.ru/
Долуханов П.М., Тимофеев В.И. Абсолютная хронология неолита Евразии (по данным радиоуглеродного датирования). Издательство "Наука", 1972.
http://www.archeologia.ru/Library/Book/6e91ff7e03d2 Портал «Археология России». http://www.archeologia.ru/
Екеев Н.В. Чоросы – Ойратские князья.
http://e-lib.gasu.ru/da/archive/2003/10/19.html, Древности Алтая №10, 2003. Электронная библиотека ГАГУ. http://e-lib.gasu.ru/
Казласов Р.Л. Государства Южной Сибири в XVI-XVIII веках. http://e-lib.gasu.ru/da/archive/2003/10/13.html Древности Алтая. №10. 2003. http://e-lib.gasu.ru/da/ Электронная библиотека ГАГУ. http://e-lib.gasu.ru/
Карафет Т.М., Зегура С.Л., Хаммер М.Ф. Историческое освоение человеком новых территорий: роль древних популяций Азии в заселении Америки. Вестник ВОГиС, 2006. Том 10, №1. С. 7-23.
http://www.bionet.nsc.ru/vogis/pict_pdf/2006/t10_1/vogis_10_1_01.pdf Институт цитологии и генетики Сибирского отделения РАН.
http://www.bionet.nsc.ru/boardtest/cgi-bin/catalog/main.pl
Колёсов А. Хинди-руси бахай-бахай с точки зрения ДНК генеалогии, или откуда есть пошли славяне. № 531, 10 июня 2007 г.
http://www.lebed.com/2007/art5034.htm\ независимый альманахhttp://www.lebed.com/
Маточкин Е.П. Архетипические образы в скифском искусстве Горного Алтая. //Итоги изучения скифской эпохи Алтая и сопредельных территорий. Барнаул. 1999. С. 122-126. Интернет-портал по истории Алтая. http://new.hist.asu.ru/
Нанзатов Б.З. К этногенезу бурят по материалам этнонимии. http://mion.isu.ru/pub/turov/3.html Сайт Иркутский мион. http://mion.isu.ru/
Носовский Г.В., Фоменко А.Т. Империя. Русь, Турция, Китай, Европа, Египет. Новая математическая хронология древности. Москва, 1995. http://chronologia.org/imperia/imperia.html
[ХРОН4] Носовский Г. В., Фоменко А.Т. Новая хронология Руси. Русь. Англия. Византия. Рим. (В трех томах). Изд. РИМИС. 2005. //chronologia.org/xpon4/index.html Сайт проекта «Новая Хронология». //chronologia.org
[ХРОН5] Носовский Г. В., Фоменко А.Т. Империя. Изд. РИМИС. 2005. //chronologia.org/xpon5/index.html Сайт проекта «Новая Хронология». //chronologia.org
Носовский Г.В., Фоменко А.Т. «Египетский альбом» (Взгляд на памятники Древнего Египта от Наполеона до Новой Хронологии) 2006, Ремис. http://chronologia.org/eg_albom1.html
Носовский Г.В., Фоменко А.Т. Казаки-арии: из Руси в Индию. АСТ-АСТРЕЛЬ, 2007.
Паршев А.П. Почему Россия не Америка.
http://www.lib.csu.ru/Discussion/003218/index.htm
Сайт Новая Хронология. //chronologia.org/
[Сайт Turkic History] Сайт Turkic History http://www.turkicworld.org/
[Тюрин, 2005, Европейцы голоцена] Тюрин А.М. Европейцы голоцена по данным радиоуглеродного датирования.
/volume3/turin_eu.html Электронный сборник статей «Новая Хронология». Выпуск 3. 2005. /volume3/ Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2005, Алгоритмы] Тюрин А.М. Алгоритмы фальсификации и ре-фальсификации результатов радиоуглеродных датировок. /volume3/turin_alg.html Электронный сборник статей «Новая Хронология». Выпуск 3. 2005. /volume3/ Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2007, Датирование, МЛП] Тюрин А.М. Датирование Малого Ледникового Периода. /volume5/tur_mlp.html Электронный сборник статей «Новая Хронология». Выпуск 5. 2007. /volume5/Сайт: Новая Хронология.
//chronologia.org/
[Тюрин, 2007, Алгоритм, Неолит] Тюрин А.М. Алгоритм радиоуглеродного датирования археологических культур неолита Евразии.
/volume5/neolit.html Электронный сборник статей «Новая Хронология». Выпуск 5. 2007. /volume5/ Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2007, Интеграция, Скифы] Тюрин А.М. Интеграция скифской археологической культуры в Новую Хронологию Фоменко и Носовского. /volume6/tur_int.html Электронный сборник статей «Новая Хронология». Выпуск 6. 2007. /volume6/index.html Сайт: Новая Хронология. //chronologia.org/
[Тюрин, 2007, Биология, Скифы] Тюрин А.М. Биологические маркеры Руси-орды и скифская археологическая культура.
/volume6/tur_biomark.html Электронный сборник статей «Новая Хронология». Выпуск 6. 2007.
/volume6/index.html Сайт: Новая Хронология. //chronologia.org/
[Этнический состав] Этнический состав населения Горного Алтая в XVII-XVIII вв. http://altay-250-letie.ru/letopis_sobitiy/dzungaria.html Сайт «Алтай» http://altay-250-letie.ru/
Bamshad M., Kivisild T., Watkins W.S., Dixon M., Ricker C.E., Rao B.B., Naidu M., Prasad B.V.R., Reddy P.G., Rasanayagam A., Pariha S.S., Villems R., Redd A.J., Hammer M.F., Nguen S.V., Carroll M.L., Batzer M.A. and Jorde L.B. Genetic Evidence on the Origins of Indian Caste Populations. Genome Res. 2001 Jun;11(6):994-1004. http://jorde-lab.genetics.utah.edu/elibrary/Bamshad_2001a.pdf Jorde Laboratory.
http://jorde-lab.genetics.utah.edu/
[Distribution of Blood] Distribution of Blood Types.
http://anthro.palomar.edu/vary/vary_3.htm MODERN HUMAN VARIATION: An Introduction to An Introduction to Contemporary Human Biological Diversity. http://anthro.palomar.edu/vary/
Cavalli-Sforza L. L. Genes, peoples, and languages. Proc. Natl. Acad. Sci. USA
Vol. 94, pp. 7719-7724, July 1997. http://www.pnas.org/cgi/content/full/94/15/7719 Сайт Proc. Natl. Acad. Sci. USA http://www.pnas.org/
