К вопросу о географии гаплогруппы Y-хромосомы I1 в Восточной Европе
Тюрин Анатолий Матвеевич, к.г.-м.н.
Ключевые слова: популяционная генетика, Y-хромосома, гаплогруппа I1, Русская равнина
Оглавление
1. Постановка задачи
2. Общие сведения о географии гаплогруппы I-М170
3. Формальная интерпретация частот гаплогрупп
4. Генетические следы миграции носителей гаплогруппы I1-М253 с севера Скандинавии на юго-восток
5. Историческая реконструкция
6. Вместо заключения
Источники информации
1. Постановка задачи
В публикации [Balanovsky, 2008] приведены результаты ДНК-тестирования русских (выборка 1228 человек) из 14 регионов европейской части России. Рассмотрены географические особенности их генетического портрета. Одна из них – наличие в распределении частот гаплогруппы I1-М253 локального максимума в районе Верхней Волги и Северной Двины (верхняя схема на рисунке 1). Автор публикации [Штрунов, 2010] за счет привлечения дополнительных ДНК-данных уточнил схему распределения частот этой гаплогруппы (нижняя схема на рисунке 1). В рамках поиска возможного объяснения феномена «локальный максимум» он рассмотрел исторические и археологические данные. Результирующий вывод звучит так. «Компактный локальный максимум частот I1 в центре Русской равнинны – следствие древнейших миграций палеолитического населения Европы, приведших к созданию верхневолжской культуры (VI – V тыс. до н.э.).». Но не рассмотрена очевидная (видимая невооруженным глазом) версия трактовка пространственного распределения частот гаплогруппы I1-М253. Это и определило наше обращение к этой проблеме.
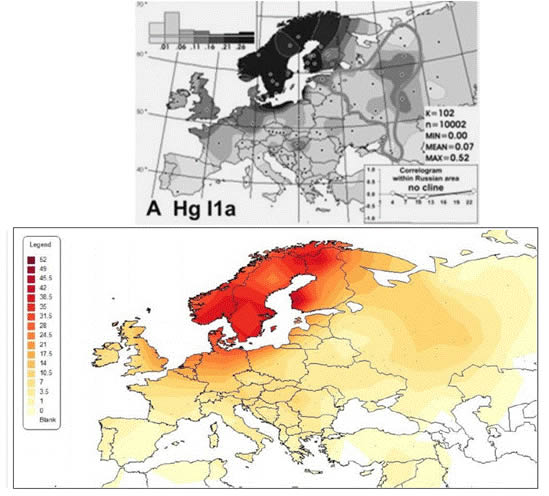
Рисунок 1. Схема частот гаплогруппы Y-хромосомы I1-М253 в Европе. Верхняя схема – [Balanovsky, 2008]. Фигурой неправильной формы обозначена область, охарактеризованная результатами исследований авторов публикации. Нижняя схема – [Штрунов, 2010].
2. Общие сведения о географии гаплогруппы I-М170
В гаплогруппе Y-хромосомы I-М170 выделяется шесть субкладов [Lechner; Rootsi, 2004]. I1-М253 (старая номенклатура – I1a) – один из них. Его максимальные частоты приурочены к Скандинавии (рисунок 1). Среди южных и северных шведов 35,7% и 26,3% носителей этого субклада [Rootsi, 2004]. Среди норвежцев – 38,9%, саамов – 28,6%. Среди финнов 28,9% носителей гаплогруппы I-М170 (без ее разделения на субклады) [Tambets, 2004]. В южном направлении частоты I1-М253 убывают: 14,8% (эстонцы – 210 человек), 4,7% (латвийцы – 86 человек) и 2,7% (белорусы – 147 человек) [Rootsi, 2004]. Частоты гаплогруппы на территории Скандинавии детально рассмотрены в публикации [Lappalainen, 2009], на территории Финляндии – в [Wiik, 2008]. Частоты гаплогруппы I1-М253 убывают от их максимума на юге Швеции в восточном, юго-восточном, северо-восточном и западном направлениях, а частоты гаплогруппы N1c (старая номенклатура – N3-TAT) – в западном направлении (рисунки 2 и 3). Особенно ярко характерное соотношение частот гаплогрупп I1-М253 и N1c проявилось на территории Финляндии (рисунок 4). Распределение частот гаплогрупп по районам Швеции и Норвегии приведено в публикации [Lappalainen, 2008, Swede] (рисунок 5). В ней учтены и данные из публикации [Dupuy, 2005]. В Норвегии выявлено 37,3% носителей субклада I1-М253, в Швеции – 38,2%. Распределение его частот на территориях стран характеризуется низкими вариациями по регионам. Причем, минимальные и максимальные частоты – 30,9% и 42,1%, выявлены в двух соседних регионах Норвегии – «западном» и «кране южном». В трех выделенных регионах Швеции частоты субклада практически стабильны – 36,3-39,8%. Повышенные частоты гаплогруппы N1c – примерно 10%, выявлены только на севере Норвегии. В публикации [Karlsson, 2006] приведены ДНК-данные по некоторым районам Швеции и Финляндии. У шведских саамов (38 человек) выявлено 31,6% носителей субклада I1-М253, 44,7% и 15,8% – гаплогрупп N1c и R1a1.
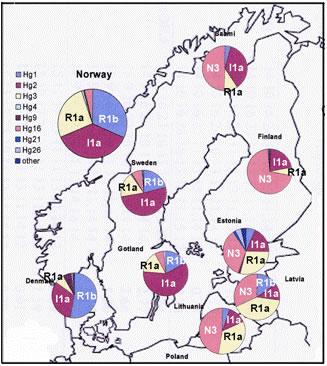
Рисунок 2. Схема частот гаплогрупп Y-хромосомы в Скандинавии и Прибалтике [Wiik, 2008] (I1a = I1-М253; N3 = N1c).
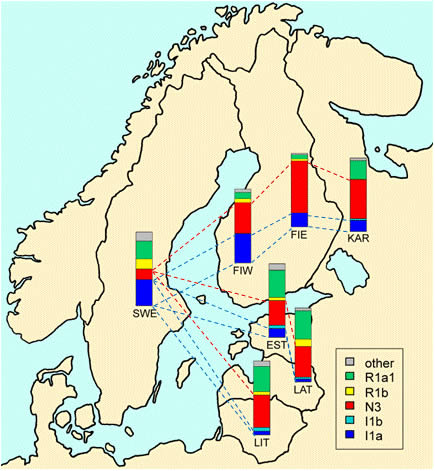
Рисунок 3. Схема частот гаплогрупп Y-хромосомы в Скандинавии и Прибалтике [Lappalainen, 2009] (I1a = I1-М253; N3 = N1c). KAR – карелы, FIE – финны восточные, FIW – финны западные, EST – эстонцы, LAT – латвийцы, LIT – литовцы, SWE – шведы. Красные и синие пунктирные линии нанесены А.М. Тюриным.
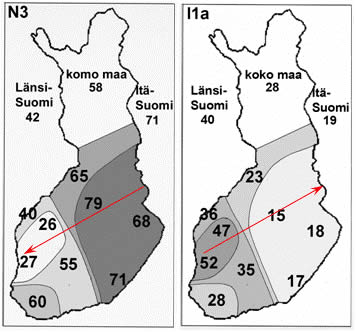
Рисунок 4. Схема частот гаплогрупп Y-хромосомы I1-М253 и N1c на территории Финляндии [Wiik, 2008] (I1a = I1-М253; N3 = N1c).
Красными стрелками показаны направления убывания частот (А.М. Тюрин).
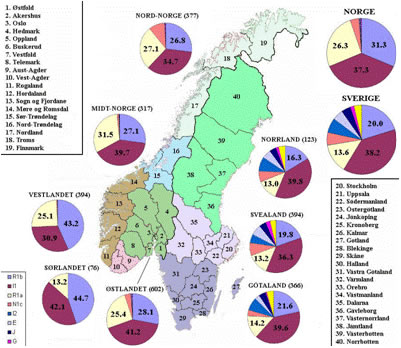
Рисунок 5. Схема частот гаплогрупп Y-хромосомы на территории Швеции и Норвегии [Lappalainen, 2008, Swede].
Выше рассмотрено распределение частот субклада I1-М253 гаплогруппы I-М170. У шведов, норвежцев и саамов практически нет носителей других ее субкладов. Среди 332 тестированных их выявлено 10 (3,0%) человек [Rootsi, 2004]. В другой выборке (305 человек) шведов выявлено 15 (4,9%) носителей субклада I2b1-M223 (старая номенклатура – I1c) [Karlsson, 2006]. Однако почти половина из них – 7 (17,0%, выборка 41 человек), проживают в провинции Vasterbotten (северная Швеция – на рисунке 5 она обозначена цифрой 39). Авторы публикации считают, что эту аномалию можно связать с территорией Германии и Дании, где наблюдается максимально высокие частоты субклада I2b1-M223 – 12,5% и 10,0% [Rootsi, 2004]). Авторы публикации [Karlsson, 2006] высказали версию о том, что жители провинции Vasterbotten, носители этого субклада, являются, в основном, потомками священников и офицеров, которые попали в нее в 17 веке из Германии и Дании. Это версия основана на системе предположений. Авторы публикации [Zerjal, 2001] выполнили анализ гаплогрупп Y-хромосомы и микросателлитных маркеров по популяциям Скандинавии и Прибалтики. Оценили степень генетических вариаций. Выделили генетические границы. Однако эти данные мы рассматривать не будем, поскольку имеется подозрение, что они устарели.
География субклада I1-М253 на территории России рассмотрена в другом разделе статьи. Из других субкладов гаплогруппы I-М170 на территории Восточной Европы наибольшие частоты имеет I2a-P37.2 (старая номенклатура – I1b). Его максимальные частоты локализованы на Балканах (боснийцы – 40,0%, хорваты – 31,2%, словенцы – 20,0%, албанцы – 17,0%) и в Северо-западном Причерноморье (гагаузы – 24,1%, молдаване – 21,1, румыны – 17,7%). Частоты субклада убывают в северном направлении. Среди эстонцев и латвийцев доля его носителей составляет 2,9% и 1,2% [Rootsi, 2004]. Среди южных русских выявлено 15,9% носителей субклада I2a-P37.2, центральных – 10,0%, северных – 5,7% [Balanovsky, 2008]. У русских выявлено 11 (0,9%) носителей субклада I2b1-M223. Кроме трех рассмотренных выше, в гаплогруппе I-М170 выделяется еще 3 субсклада, носители которых имеются в Европе [Lechner; Rootsi, 2004]. Среди русских выявлены только носители субклада I*-М170 – 7 человек (0,6%) [Balanovsky, 2008].
С анализом частот рассмотренных выше субкладов гаплогруппы I-М170 имеется одна проблема. Иногда при выполнении ДНК-исследований тестирование ограничивается идентификацией гаплогруппы без выделения ее субкладов (например [Roewer, 2008]). То есть по общему числу в выборке носителей гаплогруппы непонятно соотношение «южного» и «северного» ее субкладов. Эту проблему автор публикации [Штрунов, 2010] решил за счет идентификации субкладов по характерным для них гаплотипам. В соответствии с его комментариями, комплект данных, по которым построена карта частот субклада I1-М253 (рисунок 1), приведен в справочнике [Гаплогруппа I1 (Y-ДНК) http://ru.wikipedia.org/]. Ниже субклады гаплогруппы I-М170 мы будем называть, как и принято в большинстве публикаций, тоже гаплогруппами.
3. Формальная интерпретация частот гаплогрупп
Наша версия интерпретации локального максимума частот гаплогруппы I1-М253 в Восточной Европе проста. Никакого максимума нет. Мы имеем классический клин частот гаплогруппы I1-М253 (рисунок 1, нижний сегмент), который вытянут в юго-восточном направлении от зоны их максимума. Можно ли проверить эту версию? Да. Восточная часть максимума частот гаплогруппы I1-М253 в Скандинавии совпадает с максимумом частот гаплогруппы N1c. Если в прошлом имела место миграция жителей севера Скандинавии в юго-восточном направлении или генный дрейф, то ее (его) след сегодня будет проявляться по частотам и гаплогруппы I1-М253, и гаплогруппы N1c. В свою очередь, увеличение частот этих гаплогрупп приведет к формированию минимума частот гаплогруппы R1a. Здесь необходимо дать важные пояснения. Данные, которыми мы оперируем вслед за автором публикации [Штрунов, 2010] (данные из работ [Balanovsky, 2008; Roewer, 2008]), характеризуют в основном русских. То есть мы рассматриваем не распределение частот гаплогрупп I1-М253, N1c и R1a на северо-востоке Европы, но их распределение среди русских, проживающих в этом регионе. Доля носителей гаплогруппы R1a (R1a1) среди русских составляет примерно 50,0%. В северном направлении доля ее носителей сокращается от 56,3% (Тамбовская область) до 35,7% (Архангельская область) [Roewer, 2008]. В свою очередь в этом направлении увеличивается число носителей гаплогруппы N1c.
Итак, в соответствии с нашей версией в распределении частот гаплогрупп I1-М253, N1c и R1a на северо-востоке Европы должна быть четкая закономерность, отражающая миграцию населения или (и) дрейф генов с севера Скандинавии в юго-восточном направлении. Так ли это? Да. Ожидаемые элементы мы видим в распределении частот обозначенных гаплогрупп (рисунок 6). На них проявился клин, связанный с максимальными значениями двух первых гаплогрупп на севере Скандинавии и соответствующая ему зона пониженных частот гаплогруппы R1a. Осталось понять, чем обусловлено строго линейное направление миграции или (и) дрейфа. На рисунке 7 показана климатическая карта Европы. Принципы построения USDA-карт предложены департаментом сельского хозяйства США. Они отражают климатические зоны, ограниченные на основе параметров, определяющих вегетацию растений: минимальная температура воздуха зимой, даты первого осеннего и последнего весеннего заморозков. Клин гаплогрупп I1-М253 и N1c попадает строго в одну климатическую зону. Она примерно соответствует южной части зоны тайги.
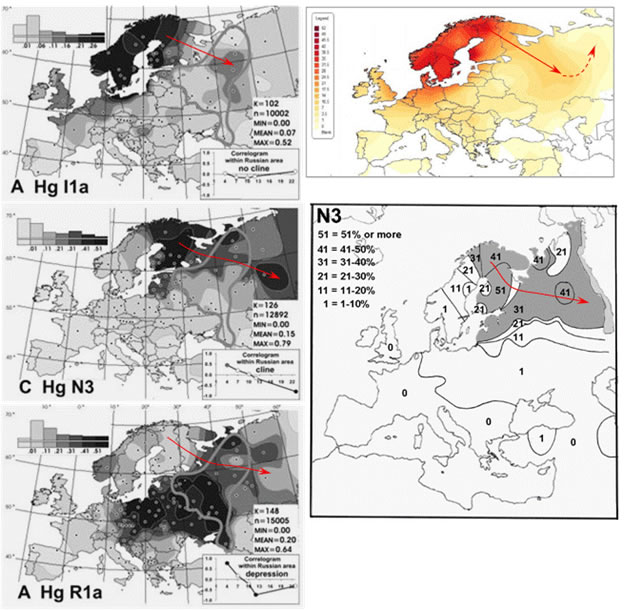
Рисунок 6. Схема частот гаплогрупп Y-хромосомы I1-М253, N1c и R1a в Европе. Три схемы слева – [Balanovsky, 2008] (I1a = I1-М253; N3 = N1c). Фигурой неправильной формы обозначена область, охарактеризованная результатами исследований авторов публикации. Верхняя схема справа – частоты гаплогруппы I1-М253 [Штрунов, 2010]. Нижняя схема справа – частоты гаплогруппы N1c [Wiik, 2008] (N3 = N1c). Красной стрелкой показано направление миграции носителей гаплогрупп I1-М253 и N1c (А.М. Тюрин).
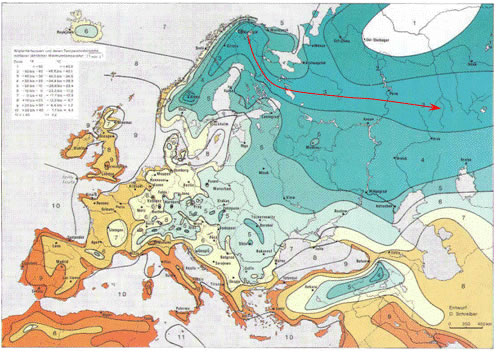
Рисунок 7. Климатическая карта (USDA) Европы [USDA-зоны]. Красной стрелкой показано направление миграции носителей гаплогрупп I1-М253 и N1c (А.М. Тюрин).
Формальная интерпретация распределения частот гаплогрупп I1-М253 и N1c на Севере Европы сводится к следующему. На севере Скандинавии в прошлом обособилось социальное сообщество, имеющее 2 отличительные характеристики. Среди ее членов доминировали носители гаплогрупп I1-М253 и N1c, а хозяйственный уклад соответствовал климатическим и природным условиям таежной зоны, предположительно ее южной части. В связи с последней особенностью, миграция членов сообщества за пределы ареала его первоначального расселения могла осуществляться только в юго-восточном направлении. Сегодня этим характеристикам соответствуют саамы (при учете их хозяйственного уклада в 19 веке), в прошлом им соответствовали их предки. Потомки мигрантов, носители гаплогрупп I1-М253 и N1c сегодня являются русскими и представителями народов Поволжья. Пока мы говорим только про Северную Европу.
Результаты нашей формальной интерпретации распределения частот I1-М253 и N1c на севере Европы имеют формальное же следствие. Сегодня большинство специалистов считает, что гаплогруппа N1c попала в Скандинавию из Восточной Европы (например, авторы публикаций [Lappalainen, 2008, Migration Waves; Wiik, 2008]). А в соответствии с нашей интерпретацией частот гаплогрупп I1-М253 и N1c, последняя попала на территорию Восточной Европы с севера Скандинавии. Если это так, то на территорию Северо-восточной Азии она попала из Восточной Европы. То есть наша интерпретация частот гаплогрупп автоматически локализует регион, переселенцы из которого являются предками носителей гаплогруппы N1c всей Северо-восточной Евразии.
Выше мы привели результаты формальной интерпретации частот гаплогрупп I1-М253 и N1c. Однако детальное рассмотрение ДНК-данных по северу Евразии привело нас к более сложной модели. Миграции было две. Одна из них произошла в то время, когда на севере Скандинавии по частоте доминировала гаплогруппа N1c, но носителей гаплогруппы I1-М253 там еще не было. Они в регионе появились позднее. Вторая волна миграции с севера Скандинавии произошла тогда, когда соотношение частот гаплогрупп N1c и I1-М253 примерно соответствовало современному. То есть генетические следы первой миграции являются как бы фоном для следов второй миграции. Данные по географии гаплогруппы N1c в Сибири позволяют проследить путь миграции. Мигранты с севера Скандинавии за Уралом продолжили движение на восток по югу зоны тайги, и дошли до Прибайкалья. Миграция происходила в одной климатической зоне – южная часть зоны тайги. От магистрального пути имелось три ответвления, по которым мигрировали отдельные группы: Минусинская котловина – Тува – Монголия; Забайкалье – Монголия; бассейн Лены. Но этот вопрос нуждается в специальном рассмотрении. Ниже детально рассмотрены следы второй миграции с севера Скандинавии.
4. Генетические следы миграции носителей гаплогруппы I1-М253 с севера Скандинавии на юго-восток
ДНК-данных по карелам мы не нашли. Судя по информации на рисунке 3 среди них порядка 15,0% носителей гаплогруппы I1-М253. По разным выборкам, характеризующим Архангельскую область, получены разные частоты интересующих нас гаплогрупп. В публикации [Balanovsky, 2008] приведены результаты тестирований в трех старых городах (сегодня это большие села) Архангельской области: Мезень (тестировано 54 человека) и Пинега (113 человек) – север области, Красноборск (91 человек) – ее юго-восток. В Мезени носителей гаплогруппы I1-М253 нет. В Пинеге выявлен 1 (0,9%) ее носитель, в Красноборске – 12,1%. Носители гаплогруппы I2a-P37.2 выявлены в Пинеге – 4,4% и Красноборске – 9,9%. Кроме того, в выборке, характеризующей последний город, выявлено 3 (3,3%) носителя гаплогруппы I*-М170. Носителей гаплогруппы N1c среди жителей городов – 46,3%, 23,7% и 36,3%, гаплогруппы R1a – 44,4%, 39,5% и 19,8%. Обращаем внимание на жителей Красноборска. Среди них носителей гаплогрупп I1-М253, N1c и R1a – 9,9%, 36,3% и 19,8%. Среди саамов (127 человек) их 25,9% (I-М170), 47,2% и 11,0% [Tambets, 2004]. Генетические портреты жителей Красноборски и саамов по рассматриваемым маркерам близки. В публикации [Roewer, 2008] приведены результаты тестирования жителей всей Архангельской области (42 человека). Среди них выявлено 31,0%, 28,6% и 35,7% носителей гаплогрупп I-М170, N1c и R1a. В публикации [Штрунов, 2010] приведена цифра из (Mirabal et al. 2009): в выборке, характеризующей жителей Архангельска выявлено 14,2% носителей гаплогруппы I1-М253. В справочнике [Гаплогруппа I1 (Y-ДНК) http://ru.wikipedia.org/] проведено еще 2 цифры: в Архангельске –7,6% (Underhill 2007), в Архангельской области – 7,0% [Roewer, 2008] носителей гаплогруппы I1-М253. С цифрой «7,0%» мы не можем согласиться. В выборке из последней публикации выявлено 31,0% носителей I-М170. Носителей гаплогруппы I1-М253 не может быть меньше 19-25%. Наши цифры основаны на простом расчете. Среди русских центральных областей (364 человека) выявлено 10,0% носителей гаплогруппы I2a-P37.2 [Balanovsky, 2008]. Кроме того, среди них выявлено 1,6% и 0,6% – носителей гаплогрупп I2b-M223 и I*-М170. При учете тенденции убывания частот I2a-P37.2 в Восточной Европе с юга на север, среди жителей Архангельской области не может быть носителей I2a-P37.2 больше 10%. Это при том, что для северных русских (380 человек) их число составляет 5,7%. В городе Вологда (121 человек) их 8,3%. Носителей гаплогруппы I2b-M223 в Вологде 1,7%. Эти цифры нужно вычесть из числа носителей гаплогруппы I-М170 выборки [Roewer, 2008]: 31,0% – 10-12,0% = 19-20,0% носителей гаплогруппы I1-М253. Это оценка минимум. Оценка максимум: 31,0% – 5,7% = 25,0%. Но достоверность этих цифр не высокая, поскольку выборка включат всего 42 человека. Тем не менее, их следует принять во внимание.
С Архангельской областью связан еще один непонятный момент. На карте частот гаплогруппы I1-М253 западнее Белого моря должно быть 3 точки: «Мезень», «Пинега» и «Красноборск». В Мезени носителей гаплогруппы I1-М253 нет. В Пинеге выявлен 1 (0,9%) ее носитель, в Красноборске – 12,1%. То есть линия нулевых частот должна «уткнуться» в Белое море немного северней Архангельска. Карта из публикации [Balanovsky, 2008] (рисунок 1) этому соответствует. Однако, точкам «Мезень» и «Пинега» не соответствует рисовка на карте из публикации [Штрунов, 2010]. Скорее всего, ее автор их не учел. Между тем, именно эти точки несут принципиальную информацию для правильной интерпретации характера распределения частот гаплогруппы I1-М253.
В соответствии с нашей гипотезой в первой волне миграции с севера Скандинавии доминировали носители гаплогруппы N1c. Сегодня их потомки примерно равномерно распределены (23,7-46,3%) по территории всей Архангельской области. Носители гаплогруппы I1-М253 в первой волне мигрантов отсутствовали. Но во второй волне миграции с северной Скандинавии носителей гаплогрупп N1c и I1-М253 было примерно поровну. Переселенцы второй волны заселили только юг Архангельской области, но не попали на ее север. Поэтому в старых городах Мезени и Пинеге сегодня практически нет носителей последней гаплогруппы. На юго-востоке области в Красноборске – их 12,1%. Скорее всего, на юго-западе области их больше 20,0%. Число носителей гаплогруппы I1-М253 в Архангельске – 14,2%, примерно соответствует их среднему числу для всей области. Возможно, эта цифра несколько выше – порядка 20,0%.
Южнее Архангельской находится Вологодская область. В характеризующей ее выборке (40 человек) 27,5%, 15,0% и 37,5% носителей гаплогрупп I-М170, N1c и R1a [Roewer, 2008]. По гаплотипам этой выборки автор публикации [Штрунов, 2010] выявил 17,0% носителей гаплогруппы I1-М253. Другая выборка (121 человек) характеризует Вологду [Balanovsky, 2008]. В ней 11,6%, 8,3% и 1,7% носителей гаплогрупп I1-М253, I2a-P37.2 и I2b-M223. Всего носители гаплогруппы I-М170 составляют 21,5%, из них доля носителей гаплогруппы I1-М253 составляет 53,5%. Эти цифры соответствует 27,5% носителей гаплогруппы I-М170 в Вологодской области. Среди них доля носителей гаплогруппы I1-М253 составляет 46,8%. Некоторое расхождение в количестве носителей I-М170 можно объяснить за счет большого процента жителей города Вологда, являющихся потомками переселенцев с юга. Число носителей гаплогрупп N1c и R1a в Вологодской области – 15,0% и 37,5% [Roewer, 2008], в Вологде – 35,5% и 33,1% [Balanovsky, 2008]. К юго-западу от Вологодской области расположены Тверская (43 человека) и Новгородская (40 человек) области. В них выявлено 5,0% и 3,0% носителей гаплогруппы I1-М253 [Гаплогруппа I1 (Y-ДНК) http://ru.wikipedia.org/]. Носителей гаплогруппы N1c – 10,6% и 27,5%. Повышенное число носителей последней гаплогруппы в Новгородской области связано с территорией Прибалтики. В соседней Эстонии их 30,6% [Tambets, 2004]. Тверская область ограничивает с юго-запада зону миграций носителей гаплогрупп N1c и I1-М253 с севера Скандинавии. Таким образом, мы четко локализовали ее географически: от северной границы Тверской области до южного окончания Белого моря.
Южнее Вологодской области имеются данные по селу Унжа Костромской области [Balanovsky, 2008]. В выборке (52 человека) выявлено 11,5%, 11,5% и 3,8% носителей гаплогрупп I1-М253, I2a-P37.2 и I2b-M223. Всего 26,8% носители гаплогруппы I-М170. Доля носителей гаплогруппы I1-М253 составляет 42,9%. Число носителей гаплогрупп N1c и R1a в Унже – 13,5% и 32,7%. В юго-восточном направлении от Костромской области ДНК-данными охарактеризованы Ивановская, Рязанская и Пензенская области. В выборке, характеризующей Ивановскую область (40 человек) 21,3%, 22,5% и 45,0% носителей гаплогрупп I-М170, N1c и R1a [Roewer, 2008]. В том числе по оценке автора публикации [Штрунов, 2010] 10,0% носителей гаплогруппы I1-М253. В Рязанской области (36 человек) 22,2%, 8,3% и 47,2% носителей гаплогрупп I-М170, N1c и R1a [Roewer, 2008]. В том числе 14,0% носителей гаплогруппы I1-М253 [Штрунов, 2010]. В Пензенской области (81 человек) 21,0%, 9,9% и 51,9% носителей гаплогрупп I-М170, N1c и R1a [Roewer, 2008]. Носителей I1-М253 – 12,5%. Эти три области являются центральной частью зоны миграции носителей гаплогруппы I1-М253 с севера Скандинавии на юго-восток.
При построении карты частот гаплогруппы I1-М253 автор публикации [Штрунов, 2010] учел точку «Татарстан» со ссылкой на атлас с сайта Генофонд.ру. Ей соответствует частота 13,0%. Такого не может быть. В выборке, характеризующей татар Поволжья (126 человек), выявлено 4,0% носителей гаплогруппы I-М170 [Tambets, 2004]. В соответствии с данными из публикации [Rootsi, 2004] (123 человека) среди татар 4,9% носителей этой гаплогруппы. Но среди них всего 1 (0,8%) носитель гаплогруппы I1-М253. Носителей гаплогрупп I2a-P37.2 и I*-М170 – 1,6% и 2,4%.
Среди коми (94 человека) и удмуртов (87 человек) носители гаплогруппы I-М170 составляют 5,3% и 1,1% [Tambets, 2004]. Но в другой выборке, характеризующей удмуртов (132 человека), выявлено 2,3% носителей этой гаплогруппы [Rootsi, 2004]. В выборке коми (110 человек) выявлено 4,5% носителей гаплогруппы I-М170, в том числе 3,6% – I1-М253 и 0,9% – I2a-P37.2. Башкиры охарактеризованы выборкой в 471 человек [Лобов, 2009]. Среди них выявлено по 1 носителю гаплогрупп I2a-P37.2 и I*-М170. У русских Башкортостана (50 человек) выявлено 4,0% носителей гаплогруппы I1-М253 [Rootsi, 2004]. Но все русские этого региона – потомки мигрантов. Среди хантов (47 человек) и ненцев (148 человек) носители гаплогруппы I-М170 не выявлены [Tambets, 2004]. То есть, за Уралом их нет. Практически их нет и у проживающих в Приуралье удмуртов. Таким образом, волна миграции с севера Скандинавии, носителей гаплогруппы I1-М253, дошла почти до Камы.
Важные выводы можно получить при рассмотрении частот гаплогруппы I2b1-M223. Их целесообразно рассматривать в абсолютном выражении и в относительном (отношение числа носителей гаплогрупп I2b1-M223 и I1-М253). У шведов, норвежцев и саамов выявлено 8 (2,4%) носителей гаплогруппы I2b1-M223 и 113 (34,0%) – I1-М253 [Rootsi, 2004]. Коэффициент отношения равен 0,071. В Мезени, Пинеге и Красноборске носителей гаплогруппы I2b1-M223 не выявлено [Balanovsky, 2008]. В Вологде их 2 (1,7%) человека, что составляет 0,147 по отношению к числу носителей гаплогруппы I1-М253. Для центральных русских коэффициент соотношения носителей этих гаплотипов составляет 0,302. Однако, для Унжи, где выявлен максимальный процент носителей гаплогруппы I1-М253 (11,5%) это соотношение равно 0,076. Для южных русских соотношение носителей этих гаплогрупп равно 0,018. Коэффициент соотношения носителей гаплогрупп I2b1-M223 и I1-М253 у немцев и датчан составляет 0,500 и 0,599 [Rootsi, 2004]. Эти цифры можно проинтерпретировать так. Среди центральных русских (кроме города Унжа) имеются значимый процент потомков носителей гаплотипа I1-М253, попавших в регион из Германии и Дании. Однако, среди южных и северных русских их немного. В принципе, можно выполнить количественные расчеты. Но мы этого делать не будем. Только отметим, что соотношение носителей гаплотипов в Вологде и Унже, попавших в центральную часть максимума частот гаплогруппы I1-М253 на Русской равнине, соответствует таковому в Скандинавии. Это прямо и непосредственно подтверждает нашу его трактовку. Количество потомков переселенцев из Германии и Дании можно оценить и другим способом. Потомков переселенцев, носителей гаплогруппы I1-М253, в России не может быть больше, чем в Белоруссии, а среди белорусов их всего 2,7% [Rootsi, 2004].
5. Историческая реконструкция
Что могло стать причиной миграции части населения севера Скандинавии, носителей гаплогруппы I1-М253, в юго-восточном направлении? Начало Малого ледникового периода, которое обычно относят к 14-15 векам. Однако мы определили другую дату этого события на Русской равнине – примерно 1560 год [Тюрин, 2007, Датирование, МЛП]. Этим рубежом и следует датировать начало миграции. Реконструкцию событий в прошлом можно выполнить на одной из карт Восточной Европы, дата составления которой максимально близка к 1560 году. Этим условиям соответствует карта А. Дженкинсона (датируется 1562 годом), варианты которой вошли в известные атласы. На рисунке 8 приведен фрагмент карты «Russiae, Moscoviae et Tartariae Descriptio» (составлена на основе карты А. Дженкинсона) из атласа «Theatrum Orbis Terrarum» (примерно 1570 год). Полностью карта приведена на сайте [Кольские карты]. Некоторые пояснения к картам, производным от карты А. Дженкинсона, даны в публикации [Осипов, 2008]. Наше общее замечания к английским картам конца 17 века сводятся к следующему. Их политическая составляющая не в полной мере соответствует общепринятому варианту русской истории этого периода. Но этот вопрос мы здесь рассматривать не будем. Просто примем обозначенную нами карту как некое отражение политической и, следовательно, этнической ситуации в регионе. На карте показана собственно RVSSIA. На юге и востоке она граничит с TARTARIA. Пограничные области последней: CRIMIA, MORDVA, NAGAI и TVMEN. На северо-западе RVSSIA граничит с LITVANIA, на западе с LIVONIA, на северо-востоке – с PERMIA, на северо-западе – с неким политическим образованием (предположительно Швеция), включающим BIARMIA, LAPPIA и CORELIA.
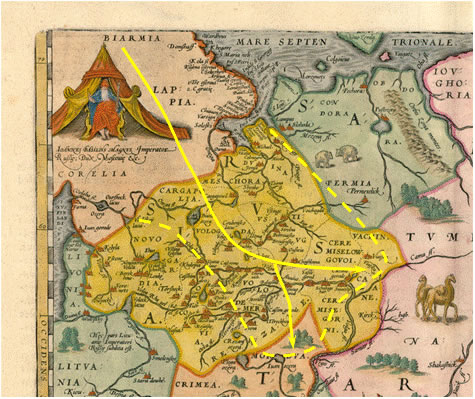
Рисунок 8. Фрагмент карты Russiae Moscoviae et Tartariae Descriptio» из атласа «Theatrum Orbis Terrarum» (примерно 1570 год) [Кольские карты].
Информация желтым цветом нанесена А.М. Тюриным. Сплошные линии со стрелками – путь миграции носителей гаплогруппы I1-М253 с севера Скандинавии.
Пунктирная линия – примерное положение границы максимальных частот гаплогруппы I1-М253 в Восточной Европе.
Нас интересует северо-западная часть карты (рисунок 8). LAPPIA – это область, где жили ЛОПари русских летописей. Это саамы. С CORELIA тоже все понятно. Зато ничего не понятно с BIARMIA. Между тем, по нашему предположению именно из региона, обозначенного этим названием, на север Русской равнины мигрировали носители гаплогруппы I1-М253. В справочнике сообщается следующее «Биармия – (Биармландия; Beormas – на англосаксонском наречии) – скандинавское название страны, ни пределы которой, ни население, ни культура, ни даже самое название до сих пор не имеет в исторической науке более или менее положительных определений. Такая неопределенность в знаниях о Б. зависит, с одной стороны, от сказочного характера саг, главного источника сведений о Б., и с другой – от полного молчания о ней наших летописцев.» [Брокгауз и Ефрон]. Но картографы 17 века прекрасно знали, где находится BIARMIA и отразили это на своих картах. Ее не нужно искать в Пермском крае. Туда создатели скандинавских саг не плавали. BIARMIA была на севере Скандинавии. До начала Малого ледникового периода климат здесь был благоприятен для существования крупных морских промыслов и ведения «лесного» хозяйства. В сагах подчеркивается богатство BIARMIA. Можно предположить, что она возникла как система колоний переселенцев с юга Скандинавии. То есть, ее населяли, в основном, потомки переселенцев, носители гаплогруппы I1-М253. Примерно в 1560 году климат резко изменился. Вести производящее хозяйство в BIARMIA стало невозможно. Ее жители, связанны с морем, переселились на юг Скандинавии. А жители, связанные с ведением «лесного» хозяйства мигрировали на юго-восток. На рисунке 8 показан путь миграции и примерное положение границы максимальных частот гаплогруппы I1-М253 в Восточной Европе. Потомками мигрантов является бОльшая часть носителей гаплогруппы I1-М253 России. Предки меньшей части ее носителей попали на территорию России из Швеции, Дании и Германии через Прибалтику. Это был самый обычный древ генов.
Город Мезень обозначен на карте (рисунок 8) как Lampas Sloboda. Переселенцы, носители гаплогруппы I1-М253, в него не попали. Среди русских жителей города ее носителей не выявлено. Восточнее и южнее Камы переселенцы тоже не попали. Среди хантов и манси нет носителей гаплогруппы I1-М253, а среди татар выявлен всего 1 (0,8%). Резкие границы зоны отсутствия носителей гаплогруппы I1-М253 в Восточной Европе как раз и свидетельствуют о недавнем ее попадании в регион. Если бы эта гаплогруппа попала в Восточную Европу тысячи лет назад, как считает автор публикации [Штрунов, 2010], то в небольшом количестве ее носители были бы среди всех популяций региона. Стрелка, обозначающая миграции в южном направлении, соответствует относительно высоким частотам носителей гаплогруппы I1-М253 в Рязанской и Пензенской областях – 14,0% и 12,5%. Скорее всего, миграция с севера Скандинавии после 1560 года, совпала с колонизацией территории Пензенской области и некоторых районов Рязанской. Юго-западная граница максимальных частот гаплогруппы I1-М253 контролируется северными границами Тверской и Новгородской областей. В них всего 5,0% и 3,0% ее носителей.
Стоит отметить некоторые особенности карты (рисунок 8), косвенно связанные с нашей реконструкцией. На севере RVSSIA показаны три региона: DVINA, MESCHORA и CARGAPOLIA. С DVINA все понятно. Это исторический регион России. Территория CARGAPOLIA и западная часть территории MESCHORA сегодня входят в республику Карелия в составе России. MESCHORA – это передача латинскими буквами слова МЕЩЁРА. Но в справочниках говорится только о племени мещёра, фигурирующем в русских летописях. Его помещают в район среднего течения Ока. На карте 1723 года («Frederik den Femtes Atlas», [Кольские карты]) показана область, подписанная двумя названиями: CARGAPOLSKAIA CARELIA и CARELIE MOSCOVITE. На востоке и западе она ограничена реками Онега и Сорока, на севере и юге – Белым морем и Онежским озером. Западнее CARELIE MOSCOVITE показана CARELIE SUEDOISE. Понятно, что слово CARGAPOLA относится к Карелии. Собственно говоря, CAR(GAP)OLA и CARELA – это одно и то же слово. ПОЛЬ – это форма слова ЭЛЬ, в данном случае оно означает «народ» или в более общей трактовке «социальная общность». G – это трансформированный суффикс АК. Но границы CARGAPOLSKAIA CARELIA на карте 1723 года включают только меньшую часть северной МЕЩЁРЫ. На карте 1570 года ее восточной границей является Двина. Можно наверняка утверждать, что в MESCHORA и CARGAPOLIA по состоянию на 1570 год и в CARGAPOLSKAIA CARELIA по состоянию на 1723 год доминировали автохтонные популяции. То есть мигранты с севера Скандинавии в период с 1560 года по конец 16 века продвигались на юго-восток вплоть до Двины по малообжитым районам, населенным популяциями, родственными им в культурном отношении. С русским населением они впервые столкнулись на территории Вологодской области.
6. Вместо заключения
По результатам геногеографического исследования сформировано две версии интерпретации распределения частот гаплогруппы I1-М253 на Русской равнине. Автор публикации [Штрунов, 2010] предполагает, что ее носители попали в регион 6000-7000 лет назад. Мы предполагаем, что это случилось 400-450 лет назад. Эти две версии верифицируются элементарно. Необходимо выполнить генохронологическое датирование общих предков кластеров гаплопипов гаплогруппы I1-М253. Как мы понимаем, соответствующая база гаплотипов у автора публикации [Штрунов, 2010] имеется. Надеемся, что он опубликует результаты своего варианта генохронологического датирования. Если этих данных пока недостаточно для выполнения достоверного датирования, то можно этот вопрос отложить на 2-3 года. Кроме того, имеется возможность выполнить по гаплотипам гаплогруппы I1-М253 геногенеалогические исследования – выявить географию распространения гаплотипов, идентичных или близких гаплотипам, характерным для Русской равнины. То есть, в решении рассмотренной в статье проблемы необходимо задействовать геногеографический, генохронологический и геногенеалогический методы исследований, которые являются составными частями популяционной генетики.
Лобов А.С.Структура генофонда субпопуляций башкир. 2009. Автореферат диссертации на соискание ученой степени кандидата биологических наук. http://ftp.anrb.ru/molgen/Lobov_AS.PDF Академическая сеть Республика Башкортостан. http://ftp.anrb.ru/
Осипов И.А. Антоний Дженкинсон и карта России 1562 года. Сыктывкар, 2008. http://biarmia.narod.ru/library/txts/osipov.html#7_ Кольские карты из глубины веков до наших дней. http://biarmia.narod.ru/
[Тюрин, 2007, Датирование, МЛП] Тюрин А.М. Датирование Малого Ледникового Периода. /volume5/tur_mlp.html Электронный сборник статей «Новая Хронология». Выпуск 5. 2007.
/volume5/ Сайт: Новая Хронология.
//chronologia.org/
[Брокгауз и Ефрон] Энциклопедия Брокгауза и Ефрона. http://gatchina3000.ru/brockhaus-and-efron-encyclopedic-dictionary/index.htm
[USDA-зоны] Что такое USDA-зоны и с чем их едят?
http://myflora.org.ua/content/articles/usda_zone/usda_zone.html MYFLORA http://myflora.org .ua/
Штрунов А. Происхождение гаплогруппы I1-М253 в Восточной. Европе. The Russian Journal of Genetic Genealogy (Русская версия), Vol 2, No 1 (2010). http://rjgg.molgen.org/index.php/RJGGRE/article/view/44 The Russian Journal of Genetic Genealogy. http://rjgg.molgen.org/
Balanovsky O., Rootsi S., Pshenichnov A., Kivisild T., Churnosov M., Evseeva I., Pocheshkhova E., Boldyreva M., Yankovsky N., Balanovska E. and Villems R. Two Sources of the Russian Partilineal Heritage in Their Eurasian Context . The American Journal of Human Genetics, Volume 82, Issue 1, 236-250, 2008. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2253976 PubMed Central. http://www.pubmedcentral.nih.gov/
Dupuy B.M., Stenersen M., Lu T.T., Olaisen B. Geographical heterogeneity of Y-chromosomal lineages in Norway. Forensic Sci Int. 2006 Dec 1;164(1):10-9. Epub 2005, Dec 7. http://vetinari.sitesled.com/norway.pdf GENETIC CHAOS. http://vetinari.sitesled.com/
Karlsson A.O., Wallerstrom T., Goherstro A., Holmlund G. Y-chromosome diversity in Sweden – A long-time perspective. European Journal of Human Genetics (2006) 14, 963–970.
http://www.nature.com/ejhg/journal/v14/n8/pdf/5201651a.pdf NATURE.COM http://www.nature.com/
[Lappalainen, 2008, Migration Waves] Lappalainen T., Laitinen V., Salmela E., Andersen P., Huoponen K., Savontaus M.-L., Lahermo P. Migration Waves to the Baltic Sea Region. Ann Hum Genet. 2008 May;72(Pt 3):337-48. Epub 2008 Feb 19. http://www.zincavage.org/Lappalainen2008.pdf WEB SITE OF DAVID ZINCAVAGE. http://www.zincavage.org/
[Lappalainen, 2008, Swede] Lappalainen T., Hannelius U., Salmela E., von Dobeln U., Lindgren C.M., Huoponen K., Savontaus M.L., Kere J., Lahermo P. Population structure in contemporary Sweden--a Y-chromosomal and mitochondrial DNA analysis. Ann Hum Genet. 2009 Jan;73(1):61-73. Epub 2008, Nov 26. http://www.ncbi.nlm.nih.gov/pubmed/19040656 NCBI. http://www.ncbi.nlm.nih.gov/
Lappalainen T. Human genetic variation in the Baltic sea region: features of population history and natural selection. Doctoral dissertation (article-based). University of Helsinki, Faculty of Biosciences, Department of Biological and Environmental Sciences, Division of Genetics Finnish Genome Center, Institute for Molecular Medicine Finland. 2009-05-15. https://oa.doria.fi/handle/10024/44624 https://oa.doria.fi/bitstream/handle/10024/44624/humangen.pdf Doria etusivu https://oa.doria.fi/
Lechner M. Learn about Y-chromosome Haplogroup I. http://www.genebase.com/tutorial/item.php?tuId=12 GENEBASE http://www.genebase.com/
Roewer L., Willuweit S., Kruger C., Marion N. Rychkov S., Morozowa I., Naumova O., Schneider Yu., Zhukova O., Stoneking M., Nasidze I. Analysis of Y chromosome STR haplotypes in the European part of Russia reveals high diversities but non-significant genetic distances between populations. International Journal of Legal Medicine, 122, №.3, 2008.
Rootsi S., Magri C., Kivisild T., Benuzzi G., Help H., Bermisheva M., Kutuev I., Barac L., Pericic M., Balanovsky O., Pshenichnov A., Dion D., Grobei M., Zhivotovsky L.A., Battaglia V., Achilli A., Al-Zahery N., Parik J., King R., Cinnioglu C., Khusnutdinova E., Rudan P., Balanovska E., Scheffrahn W., Simonescu M., Brehm A., Goncalves R., Rosa A., Moisan J.P., Chaventre A., Ferak V., Furedi S., Oefner P.J., Shen P., Beckman L., Mikerezi I., Terzic R., Primorac D., Cambon-Thomsen A., Krumina A., Torroni A., Underhill P.A., Santachiara-Benerecetti A.S., Villems R., Semino O. Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe. Am J Hum Genet. 2004 Jul;75(1):128-37. Epub 2004 May 25. http://www.familytreedna.com/pdf/DNA.RootsiHaplogroupISpread.pdf Family Tree DNA. http://www.familytreedna.com/
Tambets K., Rootsi S., Kivisild T., Help H., Serk P., Loogvali E-L., Tolk H-V., Reidla М., Metspalu E., Pliss L., Balanovsky O., Pshenichnov A., Balanovska E., Gubina M., Zhadanov S., Osipova L., Damba L., Voevoda M., Kutuev I., Bermisheva M., Khusnutdinova E., Gusar V., Grechanina E., Parik J., Pennarun E., Richard C., Chaventre A., Moisan J.-P., Barac L., Pericic M., Rudan P., Terzic R., Mikerezi I., Krumina A., Baumanis V., Koziel S., Rickards O., Stefano G., F., Anagnou N., Pappa K.I., Michalodimitrakis E., Ferak V., Furedi S., Komel R., Beckman L., and Villems R. The Western and Eastern Roots of the Saami—the Story of Genetic «Outliers» Told by Mitochondrial DNA and Y Chromosomes. Am J Hum Genet. 2004; 74(4): 661–682. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1181943 PubMed Central (PMC). http://www.pubmedcentral.nih.gov/
Wiik K. Suomen miehet. Mita Y-kromosomin DNA kertoo suomalaisten miesten alkuperasta? Tiedosto ’CD SuoMie’10.5.2008. http://www.wiik.fi/kalevi/Suomenmiehet.pdf Wiikin Juoksusivu. http://www.wiik.fi/
Zerjal T., Beckman L., Beckman G., Mikelsaar A.-V., Krumina A., Kuinskas V., Hurles M.E., Tyler-Smith C. Geographical, Linguistic, and Cultural Influences on Genetic Diversity: Y-Chromosomal Distribution in Northern European Populations. Molecular Biology and Evolution 18:1077-1087 (2001). http://mbe.oxfordjournals.org/cgi/content/full/18/6/1077 Molecular Biology and Evolution. http://mbe.oxfordjournals.org/
