Геногенеалогия евреев по гаплогруппам Y-хромосомы и аутосомным маркерам
Тюрин Анатолий Матвеевич, к.г.-м.н.
Реконструкции геногенеалогии евреев по гаплогруппам Y-хромосомы и аутосомным маркерам дают одинаковый результат: прародина евреев – Ближний Восток. Совместное рассмотрение гаплогрупп Y-хромосомы и аутосомных маркеров евреев, при учете самых общих исторических сведений, позволяет реконструировать основные этапы этногенеза по мужской линии их этнических групп и религиозных каст. Однако, результаты реконструкции одинаково хорошо соответствуют Традиционной Истории и Новой Хронологии А.Т. Фоменко и Г.В. Носовского. Как популяция, праевреи сформировались на Ближнем Востоке из коренного населения региона. Часть их потомков – мизрахи, проживали в этом регионе до середины 20 века. Другая часть праевреев начала миграцию на запад. Вначале они освоили восточную часть античной Римской Империи (средневековую Византию), потом – античную Северную Италию (средневековые торговые республики на территории современной Италии). В последнем регионе произошло смешение праевреев с местным населением. Евреи «с итальянскими корнями» мигрировали в Восточную Европу. Это положило начало формирования израэлитов ашкенази. Скорее всего, коэны (ашкенази и сефарды) сформировались на последнем этапе становления Иудаизма из мизрахов, которые прошли подготовку в религиозных центрах и были направлены в еврейские общины для выполнения ритуальных функций. Левиты ашкенази сформированы из восточнославянского субстрата. Из Северной Италии евреи мигрировали и на запад, в страны Пиренейского полуострова и Магриба. В эти же регионы была миграция и мизрахов. На основе этих двух потоков мигрантов сформировались сефарды и марокканские евреи. Относительно небольшие группы мизрахов попали на территории Эфиопии, Индии и Йемена, где создали из местного населения иудейские общины. Потомками этих иудеев являются эфиопские, индийские и йеменские евреи.
Ключевые слова: евреи, популяционная генетика, генгенеалогия.
Оглавление
1. Введение
2. Структура социального сообщества «Евреи»
3. Гаплогруппы Y-хромосомы
4. Аутосомные маркеры
5. Обобщение результатов
6. Заключение
Источники информации
1. Введение
В последние годы форсированными темпами развиваются генетические исследования Человека. Применительно к изучению его прошлого обособились три дисциплины: геногенеалогия, геногеография и генохронология, которые являются составными частями популяционной генетики. Они базируются на одном и том же массиве данных – генетических маркерах человека. Имеются маркеры, не влияющие на биологические особенности людей, которые передаются только от отца к сыну (их выделяют в Y-хромосоме) или от матери к дочери (их выделяют в мтДНК). В Y-хромосоме выделяются два вида маркеров. Они определяют «гаплогруппу» и «гаплотип» людей. Но гаплогруппа не является совокупностью гаплотипов. Это самостоятельный маркер. Маркеры Y-хромосомы и мтДНК характеризуют происхождение человека, их носителя. Популяцию характеризует совокупность маркеров (генетический портрет), сформированная по выборке людей. Аутосомные маркеры человека передаются и от отца, и от матери. Поэтому маркеры одного человека характеризуют популяцию с относительно длительной историей. Маркеры выборки людей из популяции в своей совокупности формируют ее генетический портрет. Данные по маркерам мтДНК, характеризующие евреев по женской линии, имеются. Но они пока не дают устойчивой картины.
Способ передачи гаплогрупп Y-хромосомы и аутосомных маркеров определяет одно принципиальное отличие сформированных на их основе генетических портретов популяций. Частоты гаплогруппы Y-хромосомы характеризуют «древнюю» геногенеалогию популяций, по которой возможна локализация их прародины по мужской линии. Аутосомные маркеры в своей совокупности характеризуют «недавнюю» геногенеалогию популяций. По ним можно локализовать их прародину в недалеком прошлом.
Основной метод геногенеалогии сводится к формированию генетического портрета изучаемой популяции и его сопоставлению с генетическими портретами других популяций. «Генетическая дистанция» является мерой сходства между популяциями.
Генетический портрет евреев, отраженный в гаплогруппах и гаплотипах Y-хромосомы, рассмотрен в наших статьях [Тюрин, 2010, Скорости мутаций; Тюрин, 2010, Датирование евреев]. При этом основное внимание уделено генохронологическому аспекту. Результаты первого широкомасштабного исследования аутосомных маркеров 7 популяций евреев приведены в статье [Atzmon, 2010]. Она опубликована в журнале «The American Journal of Human Genetics» 3 июня 2010 года. Через несколько дней – 9 июня 2010 года, опубликована наша статья [Тюрин, 2010, Генетика евреев], в корой критически рассмотрены фактические данные, приведенные в публикации [Atzmon, 2010]. В этот же день в журнале «Nature» опубликована статья [Behar, 2010], в которой рассмотрены аутосомные маркеры 14 популяций евреев. В августе опубликована статья А.А. Клесова, в которой он критически рассмотрел фактические данные из публикаций [Atzmon, 2010; Behar, 2010]. Еще одна статья, в которой рассматривается генетика евреев [Zoossmann-Diskin, 2010], опубликована 6 октября в журнале «Biology direct». В статье рассмотрены аутосомные маркеры 5 популяций евреев. Рассмотрены также генетические маркеры Y-хромосомы и мтДНК. Представлялось целесообразным рассмотреть геногенеалогию евреев совместно по гаплогруппам Y-хромосомы и аутосомным маркерам.
2. Структура социального сообщества «Евреи»
Ведем понятие «социальное сообщество». Это группа людей, осознающих свое единство, хотя бы по одному идентификационному признаку. Все люди, входящие в социальное сообщество «Евреи», осознают свое единство хотя бы по одному признаку – они считают себя евреями (иудеями). Этого и достаточно. Среди евреев выделяется несколько этнических групп. Это, прежде всего, ашкенази, сефарды и мизрахи. В 16 веке ашкенази жили в Европе от Луары на западе, до Днепра на востоке и от Рима на юге до Дании на севере. Но наибольшая их часть проживала на территориях Польши, Белоруссии, Литвы и Западной Украины. Сефарды – это евреи Пиренейского полуострова. Они вынуждены были покинуть его в 1492-1496 годах. Мизрахи – евреи, жившие на территории Сирии, Ливана, Ирака и Ирана до середины 20 века. Другие этнические группы тоже четко обособлены. Это евреи Северной Африки (Туниса, Алжира, Марокко), Йемена, Грузии, Курдистана, Индии, Эфиопии, горские евреи Дагестана, караимы Крыма и другие. Особенностью евреев является наличие у них трех каст: израэлиты, коэны и левиты. Две последние касты являются жреческими. В соответствии с этнической, географической и кастовой структурой евреев, структурируются и характеризующие их ДНК-данные.
3. Гаплогруппы Y-хромосомы
Фактические данные по гаплогруппам и гаплотипам Y-хромосомы евреев приведены в публикациях [Adams, 2008; Behar, 2003; Hammer, 2009]. В обобщенном виде они рассмотрены в статьях [Тюрин, 2010, Скорости мутаций; Тюрин, 2010, Датирование евреев].
Наиболее представительные ДНК-данные по евреям приведены в публикации [Hammer, 2009]. Они включают гаплогруппы и гаплотипы 1575 евреев, в том числе 215 коэнов, 154 левитов, 738 израэлитов и 468, не знающих к какой из трех каст они относятся. Кроме того были тестированы представители нееврейских популяций (всего 2099 человек) Европы, Индии, Ближнего Востока, Северной Африки и Центральной Азии. Данные по частотам гаплогрупп в табличной форме приведены только для коэнов и израэлитов без разделения их на ашкенази и не ашкенази. Среди них доминируют носители гаплогруппы J. Среди коэнов их 75,3%, израэлитов – 36,1%. Гаплогруппа J делится на два субклада J1 и J2. Среди коэнов их носители составляют 46,0% и 29,3%, среди израэлитов – 15,1% и 21,1%. Все коэны, носители субклада J1, имеют его линию J-P58*. Ее можно записать и так J-P58(xM367, M368, M369) или J1e*. Среди коэнов ашкенази 51,6% ее носителей, среди коэнов не ашкенази – 38,7%. Среди носителей линии J-P58* выделяется модальный галотип, который получил название «модальный гаплотип коэнов» (Cohan Modal Haplotype – CMH). Среди евреев, носителей гаплогруппы J2 для линий J-M410*, J-M12 и J-M318 выделяются свои модальные гаплотипы. Выделен модальный гаплотип и для коэнов, носителей линии R-M269 субклада R1b. Вся вышеперечисленная номенклатура соответствует Y-DNA Haplogroup Tree 2009 [http://www.isogg.org/tree/index.html].
Гаплогруппа J характерна для популяций Ближнего Востока [Hammer, 2009]. У них доминирует та же ее линия, что и у евреев – J-P58*. Среди бедуинов Аравийского полуострова (выборка 34 человека) выявлено 64,7% ее носителей, среди друзов (329 человек) – 15,8%, иракцев (29 человек) – 24,1%, иорданцев (179 человек) – 55,3%, ливанцев (22 человека) – 18,2%, палестинцев (34 человека) – 32,9%, саудовских арабов (20 человека) – 40,0%, сирийцев (84 человека) – 23,8%, йеменцев (15 человек) – 66,7%. Однако среди тестированных представителей этих популяций не выявлены носители 17 маркерного варианта гаплотипа CMH. Этот гаплотип вообще отсутствует у не евреев. К распространенным среди евреев относятся гаплогруппы E, G и Q. Среди коэнов их 4,7%, 3,6% и 0,95%. среди израэлитов – 13,8%, 9,8% и 4,7%. Распространены и линии гаплогруппы R. Среди коэнов имеется 2,3% носителей линии R1a1(M17) и 5,6% – R1b1b2(M269). Среди израэлитов их 4,2% и 8,0%.
Таким образом, ДНК-данные, характеризующие коэнов, имею ярко выраженные отличительные особенности. Это доминирование среди них носителей гаплогруппы J (75,3%) и ее линии J-P58* (51,6% среди коэнов ашкенази и 38,7% – не ашкенази). Причем, 64,6% коэнов, носителей линии J-P58*, имеют 17 маркерный CMH, характерный только для евреев. Эти же особенности проявляются среди израэлитов. Среди них 36,1% носителей гаплогруппы J и 14,5% ее линии J-P58*. Среди последних имеется 10,7% носителей CMH. У левитов выявлено 13,0% носителей линии J-P58*, но среди них нет носителей CMH.
Другой массив ДНК-данных, характеризующий евреев, приведен в публикации [Behar, 2003]. Они имеют невысокое разрешение. Выборка включает коэнов ашкенази (76 человек), левитов ашкенази (60 человек), израэлитов ашкенази (100 человек), коэнов сефардов (69 человек), левитов сефардов (31 человек) и израэлитов сефардов (63 человека). Кроме того для сопоставления приведены данные по немцам (выборка 88 человек), норвежцам (83 человека), лужицким сербам (112 человек) и белорусам (360 человек). В этом абзаце рассмотрены только ашкенази. Число носителей гаплогруппы J у коэнов составило 75,4%, у левитов – 10,0%, израэлитов – 37,0%. Однако оказалось, что число носителей гаплогруппы R1a1 у левитов – 51,7%, соответствует сербам (63,4%) и белорусам (51,0%). Среди поляков, белорусов, украинцев и русских (за исключением проживающий в северных областях Европы) число носителей этого маркера немногим больше 50,0%. В поле двух главных компонент четко обособилось три кластера – все евреи, кроме левитов ашкенази; немцы и норвежцы; белорусы, сербы и левиты ашкенази. Среди левитов ашкенази, носителей гаплогруппы R1a1, выделился модальный гаплотип. В его кластер попало 74,0% ее носителей или 38,0% всех левитов. Это для 6 маркерных гаплотипов. Для 12 маркерных гаплотипов частота модального гаплотипа в линии R1a1 составила 58,0%. У коэнов и израэлитов число носителей гаплогруппы R1a1 - 1,3 и 4,0%. Число носителей гаплогруппы E*(xE3a) у коэнов, левитов и израэлитов – 4,0%, 20,0% и 22,0%.
Авторы публикации [Adams, 2008] выполнили сопоставление генетического портретов сефардов (выборка 174 человека), жителей Пиренейского полуострова (1140 человек) и жителей стран Магриба (361 человек). У жителей Пиренейского полуострова доминируют носители линии R1b3* (54,6%). У жителей Магриба – E3b2 (54,3%). По мнению авторов публикации, на полуострове имелось взаимное влияние трех популяций: его коренных жителей (испанцев, португальцев, басков), арабов Магриба и сефардов. Коренные жители «обменялись» с сефардами маркерами R1b3* и J. Среди первых 8,9% носителей гаплогруппы J, среди вторых – 11,4% линии R1b3*. От арабов Магриба коренные жители полуострова получили 4,3% носителей маркера E3b2. Сефарды его практически не получили. Среди них выявлен всего 1 (0,6%) его носитель. Всего среди сефардов 9,2% носителей гапогруппы E. Среди них выявлено 15,5% носителей гаплогруппы G, а также 2,3% и 4,6% гаплогрупп Q(xQ3)* и R1a1. Среди коренных жителей Пиренейского полуострова число носителей двух последних маркеров составляет 1,2% и 0,2% соответственно. Носителей линии R1b1b2(M269) среди сефардов – 5,3%.
Данные по еще одной выборке сефардов приведены в публикации [Behar, 2003]. Наибольшее число носителей гаплогруппы J отмечено у коэнов (выборка 69 человек) – 75,4%. У левитов (31 человек) и израэлитов (63 человека) – 32,3% и 19,1%. Число носителей гаплогруппы R1a1 у каст сефардов составляет 5,8%, 3,2% и 1,6% соответственно. Число носителей гаплогруппы E*(xE3a) – 4,4%, 9,7% и 19,1%.
Таким образом, генетические портреты ашкенази и сефардов имеют яркие отличительные особенности по отношению к генетическим портретам коренных жителей регионов Европы, где они проживали до середины 20 века. Ашкенази (кроме левитов) и сефарды отличаются от коренных жителей наличием гаплогрупп, которые сегодня доминируют по частотам в популяциях Ближнего Востока. От последних евреи отличаются гаплогруппами, которые они получили от популяций Европы, и гаплотипами ближневосточных гаплогрупп. Частоты ближневосточных и европейских маркеров в кастах ашкенази различны. Наибольшее число носителей ближневосточных маркеров у коэнов, наименьшее – у левитов. У последних доминируют частоты той же гаплогруппы, частоты которой доминируют у восточных славян. Вывод здесь однозначен. Прародиной ашкенази (кроме левитов) и сефардов является Ближний Восток. Прародиной левитов – Восточная Европа.
Вывод о том, что прародиной ашкенази является Ближний Восток, сегодня является общепринятым среди специалистов, оперирующих ДНК-данными. Его можно проиллюстрировать результатами многомерного шкалирования большого массива фактических ДНК-данных (сформирован по опубликованным данным), характеризующих популяции Евразии и Северной Африки, [Zoossmann-Diskin, 2010] (рис. 1). Восточно-европейские евреи (ашкенази) попали в один кластер (на рисунке он обведен красным эллипсом) с палестинцами, египтянами, сирийцами, киприотами, ливанцами, иорданцами, иракцами и турками. В это же кластер попали марокканские, ливанские и йеменские евреи. Обособились эфиопские, индийские, иракские и тунисские (Djerban Jews) евреи. Вместе с тем точка, характеризующая восточно-европейских евреев, попала на линию (на рисунке – красная пунктирная линия), которая отделяет европейские популяции (включая турок) от популяций Азии и Африки. По генетическому портрету, ближайшими к восточно-европейским евреям являются турки, итальянцы, греки и албанцы.
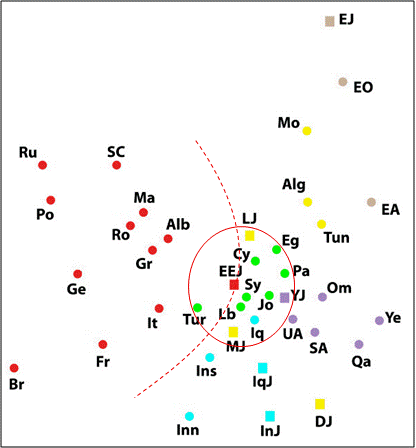
A multidimensional scaling plot of the Y-chromosomal genetic distance matrix. Stress = 0.133. Populations names are: EEJ - Eastern European Jews, IqJ - Iraqi Jews, InJ - Iranian Jews, MJ - Moroccan Jews, LJ - Libyan Jews, DJ - Djerban Jews, YJ - Yemenite Jews, EJ - Ethiopian Jews, Pa - Palestinians, It - Italians, Fr - French, Br - British, Ge - Germans, Ru - Russians, Po - Poles, SC - Serbo-Croats, Alb - Albanians, Gr - Greeks, Ma - Macedonians, Ro - Romanians, Tur - Turks, Inn - Iranians-North, Ins - Iranians-South, Iq - Iraqis, Cy - Cypriots, Sy - Syrians, Lb - Lebanese, Jo - Jordanians, SA - Saudi-Arabians, Qa - Qataris, UA - United Arab Emirates, Om - Omanis, Ye - Yemenites, Eg - Egyptians, Mo - Moroccans, Alg - Algerians, Tun - Tunisians, EO - Ethiopians Oromo, EA - Ethiopians Amhara. Squares represent Jews and circles non-Jews. Colour indicates geographic region: red - Europe, green - Eastern Mediterranean, blue - Iran-Iraq, purpule - Arabian peninsula, yellow - North-Africa, brown - Ethiopia.
Рис. 1 – Результаты обработки методом многомерного шкалирования генетических дистанций между популяциями, рассчитанных по совокупностям характеризующих их гаплогрупп Y-хромосомы [Zoossmann-Diskin, 2010]. Информация красным цветом нанесена А.М. Тюриным.
4. Аутосомные маркеры
Авторы публикации [Atzmon, 2010] привели результаты полногеномного анализа однонуклеотидного полиморфизма 7 еврейских популяций. Их выделение выполнено по географическому, этническому и этно-географическому принципам. По географическому принципу выделены итальянские, турецкие и греческие евреи. По этническому – евреи ашкенази. По этно-географическому – евреи мизрахи отдельно для Сирии, Ирака и Ирана. Всего во внимание принято 237 результатов тестирования. При их интерпретации использованы базы данных «Human Genome Diversity Project» и «The Population Reference Sample». В них включены «146 non-Jewish Middle Easterners (Druze, Bedouin and Palestinian), 30 northern Africans (Mozabite from Algeria), 1547 Europeans, and 653 individuals from other African, Asian, Latin American, and Oceanian populations.». Генетические дистанции между популяциями обработаны методами главных компонент (principal component analyses) и многомерного шкалирования (multi-dimensional scaling).
Результаты нашей интерпретации фактических данных из публикации [Atzmon, 2010] приведены в статье [Тюрин, 2010, Генетика евреев]. Вывод однозначен. Прародина евреев локализована территориями Израиля, Ливана и Сирии. Наиболее близким к их предкам является геном сирийских евреев, друзов, бедуинов и части палестинцев. Близким – турецких и греческих евреев. Часть иракских евреев генетически близка к сирийским евреям. Возможно, часть Ирака входила в регион прародины евреев. Возможно, часть предков иракских евреев являлась выходцами из Сирии.
Методология авторов публикации [Behar, 2010] практически не отличается от вышеописанной. Они рассмотрели новые результаты тестирования 121 еврея из 14 популяций совместно с результатами тестирования 1161 не евреев из 69 популяций. Первый их вывод вполне определенный. Эфиопские и индийские (Mumbai и Cochini Jews) евреи не имеют ничего общего с другими популяциями евреев и генетически неотличимы от не евреев региона их проживания. На рисунке 2 приведена выкопировка из Figure 1 рассматриваемой публикации, на которой показаны результаты обработки методом главных компонент генетических дистанций между популяциями Старого Света. В один «еврейский» кластер попали ашкенази, сефарды, сефарды Белмонта (Португалия), иракские, иранские, грузинские, азербайджанские, марроканские, узбекские евреи, а также сирийцы, самаритяне, друзы и армяне. Вблизи границ кластера попали точки, характеризующие тосканцев (Tus), испанцев (Spa), армян и грузин. Часть сирийцев и друзов в кластер не попали. Йеменские евреи попали в поле саудовских арабов.
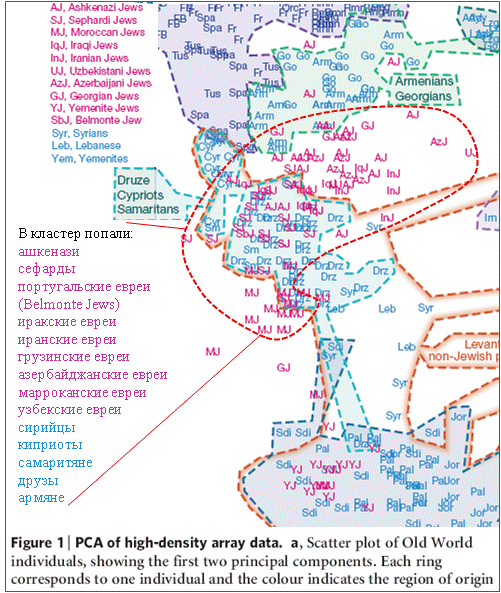
Рис. 2 – Результаты обработки методом главных компонент генетических дистанций между популяциями Старого Света, рассчитанными по совокупностям характеризующих их аутосомных маркеров [Behar, 2010]. Информация красным цветом нанесена А.М. Тюриным.
При включении в выборку только популяций западной Евразии произошло их более четкое обособление (рис. 3). Русские проявились как популяция с высокой генетической однородностью. На минимальных генетических дистанциях от них находятся генетические портреты латвийцев и белорусов. Несколько дальше – чувашей и венгров. Баски и сардинцы обособились от других популяций. Генетические портреты французов, венгров и румын находятся друг от друга на минимальных дистанциях. От французов на минимальной дистанции находятся испанцы и жители Оркнейских островов (Orcadians). Генетический портрет последних имеет принципиальную особенность. Он попадает в тренд: русские-белорусы-оркнейцы. Отметим, что именно среди жителей этих островов и севера Шотландии аномально высок (для Западной Европы) процент носителей гаплогруппы R1a1, которая доминирует у русских, белорусов, украинцев и поляков.

Рис. 3 – Результаты обработки методом главных компонент генетических дистанций между популяциями Западной Евразии, рассчитанных по совокупностям характеризующих их аутосомных маркеров [Behar, 2010]. Информация красным и желтым цветом нанесена А.М. Тюриным.
Популяция адыгейцев разделена на две субпопуляции с разными генетическими портретами. На генетический портрет одной из них «наложились» лезгины и обособленная субпопуляция турок. Эта ситуация интерпретируется достаточно просто. Предками части турок были выходцы с Северного Кавказа – адыгейцы или (и) лезгины. Скорее всего, здесь мы имеем дело со следами исхода народов Северного Кавказа в Турцию в 19 веке.
Генетический портрет турок достаточно компактный. На него частично наложились портреты субпопуляций румын. Это, скорее всего, потомки турок, которые жили в Трансильвании. Генетические портреты турок, сирийцев и иранцев находятся на минимальных генетических дистанциях и частично перекрываются. Генетический портрет киприотов находится на минимальной дистанции от генетического портрета сирийцев, но достаточно далеко от турок. Популяции грузин и армян не имеют единого генетического портрета. Армяне, в основном, попали в пограничную область между сирийцами, турками и иранцами. Грузины, в основном, попали в генетическую область иранцев.
Генетический портрет бедуинов, наименее локализованный из представленных на диаграмме. В нем выделяется три субпопуляции. Две из них накладываются на генетический портрет саудовских арабов. На него же накладываются генетические портреты палестинцев, ливанцев и частично, самаритян. Генетические портреты этих популяций накладываются и на портрет сирийцев.
По этим результатам решен вопрос йеменских евреев. Их генетический портрет попал в область саудовских арабов. По совокупности гаплогрупп Y-хромосомы йеменские евреи попали в один кластер с восточноевропейскими евреями (рис. 1). Но это не совсем так. Характеризующая их точка попала между точками, характеризующими иорданцев, палестинцев, арабов Омана и Объединенных Арабских Эмиратов. То есть, йеменские евреи – это потомки арабов, принявших иудаизм. «Еврейский» кластер разделился на два кластера. В один попали те евреи, которых называют мизрахами. Иракские евреи попали в поле сирийцев. Рядом с их областью локализовались точки, характеризующие иранских, грузинских и азербайджанских евреев. Генетически они неразличимы. В положении их общего портрета имеется яркая особенность. Он находится в «зазоре» между портретами сирийцев и иранцев и рядом с границей области турок. Почти в эту же область попала одна из субпопуляций армян.
Генетический портрет ашкенази занимает промежуточное положение между европейцами и популяциями Ближнего Востока. Причем, одна их субпопуляции частично наложалась на область румын и находится в непосредмственной близость от области тосканцев. Генетические портрет сефардов попал в область портрета ашкенази. Рядом, частично перекрываясь с ними, находится генетический портрет марокканский евреев. Генетические портреты сефардов и марокканских евреев можно объединить. Можно пойти дальше и объединить генетические портреты сефардов, марокканских евреев, ашкенази и тосканцев. В этот кластер попадает и часть киприотов.
Автор публикации [Zoossmann-Diskin, 2010] выполнил анализ аутосомных маркеров восточноевропейских, иракских, иранских, марокканских, йеменских и эфиопских евреев, а также семи европейских популяций, турок и палестинцев. Точка, характеризующая восточноевропейских евреев, попала в область европейских популяций (включая турок) и находится в непосредственной близости от точки, характеризующих итальянцев. Напомним, что в выборке [Behar, 2010] (рис. 3) тосканцы попали в «еврейский» кластер ашкенази и сефардов. В кластер генетически близких к евреям популяций (красный овал) попали турки и греки.
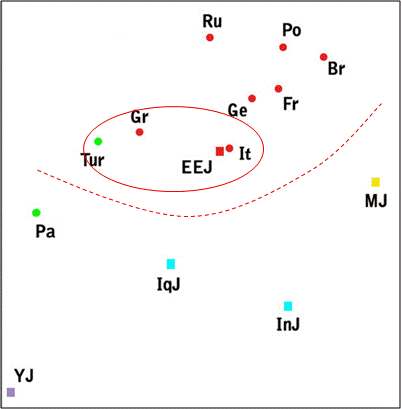
Figure 1
A multidimensional scaling plot of the autosomal genetic distance matrix excluding Ethiopian Jews. Stress = 0.100. Populations names are: EEJ - Eastern European Jews, IqJ - Iraqi Jews, InJ - Iranian Jews, MJ - Moroccan Jews, YJ - Yemenite Jews, Pa - Palestinians, Tur - Turks, Gr - Greeks, It - Italians, Ge - Germans, Br - British, Fr - French, Ru - Russians, Po - Poles. Squares represent Jews and circles non-Jews. Colour indicates geographic region: red - Europe, green - Eastern Mediterranean, blue - Iran-Iraq, purpule - Arabian peninsula, yellow - North-Africa.
Рис. 4 – Результаты обработки методом многомерного шкалирования генетических дистанций между популяциями, рассчитанных по совокупностям характеризующих их аутосомных маркеров [Zoossmann-Diskin, 2010]. Информация красным цветом нанесена А.М. Тюриным.
5. Обобщение результатов
Фактические ДНК-данные, характеризующие евреев, и результаты их обработки можно обобщить следующим образом.
1. По гаплогруппам Y-хромосомы и аутосомным маркерам популяции евреев разделились на три группы. Первая - мизрахи (сирийские, ливанские, иракские, иранские, грузинские, азербайджанские и узбекские евреи). Вторая группа включает ашкенази, сефардов и марокканских евреев. К третьей относятся эфиопские, индийские и йеменские евреи.
2. Мизрахи по генетическому портрету практически неотличимы от популяций Ближнего Востока и Ирана, причем, характеризующие их точки занимают центральное положение в совокупности генетических портретов популяций региона.
3. Ашкенази, сефарды и марокканские евреи занимают промежуточное положение между популяциями Ближнего Востока и Европы (включая турок). Сефарды, марокканские евреи и часть ашкенази ближе к ближневосточным популяциям, часть ашкенази – к европейским.
4. Эфиопские, индийские и йеменские евреи практически неотличимы от популяций коренного населения регионов их проживания. Генетический портрет индийских и йеменских евреев кардинально отличается от портрета мизрахов. Генетический портрет йеменских евреев ему соответствует так же, как портрет иорданцев, палестинцев, арабов Омана и Объединенных Арабских Эмиратов.
5. По гаплогруппам Y-хромосомы и аутосомным маркерам ашкенази по совокупности генетических дистанций находятся между популяциями Ближнего Востока и Европы (включая турок). Но имеется одно принципиальное отличие. По гаплогруппам Y-хромосомы ашкенази ближе к популяциям Ближнего Востока, а по аутосомным маркерам – к популяциям Европы. Из европейцев ашкенази наиболее близки к итальянцам. Близки к туркам и грекам.
6. Касты ашкенази и сефардов различаются по частотам ближневосточных и европейских гаплогрупп Y-хромосомы. Наибольшие частоты ближневосточных гаплогрупп у коэнов ашкенази, наименьшее – у левитов. У левитов ашкенази по частотам доминируют гаплогруппа, характерная для восточных славян. У сефардов наибольшие частоты ближневосточных гаплогрупп у коэнов, наименьшие – у израэлитов.
Реконструкция геногенеалогии евреев по гаплогруппам Y-хромосомы и аутосомным маркерам не отличается от представлений Традиционной Истории по этому вопросу. Как популяция, праевреи сформировались на Ближнем Востоке из коренного населения региона. Часть их потомков – мизрахи, проживали в этом регионе до середины 20 века. Другая часть праевреев начала миграцию на запад. Вначале они освоили восточную часть Римской Империи (территории современных Турции и Греции), потом – Северную Италию. В последнем регионе произошло смешение праевреев с местным населением. Евреи «с итальянскими корнями» мигрировали в Восточную Европу (территории современных Польши, Белоруссии, Литвы и Украины западнее Днепра). Это положило начало формирования израэлитов ашкенази. Скорее всего, коэны (ашкенази и сефарды) сформировались на последнем этапе становления Иудаизма из мизрахов, которые прошли подготовку в религиозных центрах и были направлены в еврейские общины для выполнения ритуальных функций. Левиты ашкенази сформированы из восточнославянского субстрата.
Из Северной Италии евреи мигрировали и на запад, в страны Пиренейского полуострова и Магриба. В эти же регионы была миграция и мизрахов. На основе этих двух потоков мигрантов сформировались сефарды и марокканские евреи. Относительно небольшие группы мизрахов попали на территории Эфиопии, Индии и Йемена, где создали из местного населения иудейские общины. Потомками этих иудеев являются эфиопские, индийские и йеменские евреи.
Однако по фактическим ДНК-данным, характеризующим евреев, можно выполнить реконструкцию их геногенеалогии и в рамках Новой Хронологии А.Т. Фоменко и Г.В. Носовского [Сайт Новая Хронология]. Для этого «восточную часть Римской Империи» нужно заменить на «Византия», а «Северную Италию» конкретизировать – «Торговые республики на территории современной Италии – Венеция, Генуя, Флоренция и другие».
6. Заключение
Реконструкции геногенеалогии евреев по гаплогруппам Y-хромосомы и аутосомным маркерам дают одинаковый результат: прародина евреев – Ближний Восток. Совместное рассмотрение гаплогрупп Y-хромосомы и аутосомных маркеров евреев, при учете самых общих исторических сведений, позволяет реконструировать основные этапы этногенеза по мужской линии их этнических групп и религиозных каст. Однако, результаты реконструкции одинаково хорошо соответствуют Традиционной Истории и Новой Хронологии.
Источники информации- [Сайт Новая Хронология] Сайт «Новая Хронология». //chronologia.org/
- [Тюрин, 2010, Скорости мутаций] Тюрин А.М. «Трансмиссионные», «эволюционные» и «сконструированные» скорости мутаций локусов гаплотипов Y-хромосомы. /volume10/turin_mutation.php Электронный сборник статей «Новая Хронология». Выпуск 10. 2010. /volume10/ Сайт: Новая Хронология.//chronologia.org/
- [Тюрин, 2010, Датирование евреев] Тюрин А.М. Генохронологическое датирование евреев в контексте Традиционной Истории и Новой Хронологии. /volume10/turin_datirovanie_evreev.php Электронный сборник статей «Новая Хронология». Выпуск 10. 2010.
/volume10/ Сайт: Новая Хронология. //chronologia.org/ - [Тюрин, 2010, Генетика евреев] Тюрин А.М. Молекулярная генетика евреев. http://supernovum.ru/public/index.php?doc=159 Supernovum.ru. http://supernovum.ru Электронный альманах Арт&Факт №1(13), 2010. http://artifact.org.ru/hronologiya-s-raznih-tochek/a-m-tyurin-molekulyarnaya-genetika-evreev.html Сайт Арт&Факт. http://artifact.org.ru/
- Adams S.M., Bosch E., Balaresque P.L., Ballereau S.J., Lee A.C., Arroyo E., López-Parra A.M., Aler M., Grifo M.S.G., Brion M., Carracedo A., Lavinha J., Martínez-Jarreta B., Quintana-Murci L., Picornell A., Ramon M., Skorecki K., Behar D.M., Calafell F. and Jobling M.A. The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula. The American Journal of Human Genetics, Volume 83, Issue 6, 725-736, 04 December 2008.
https://lra.le.ac.uk/bitstream/2381/4116/1/Adams08.AJHG_LRA.pdf University of Leicester. https://lra.le.ac.uk/ - Atzmon G., Hao L., Pe'er I., Velez C., Pearlman A., Palamara P.F., Morrow B., Friedman E., Oddoux C., Burns E., Ostrer H. Abraham's Children in the Genome Era: Major Jewish Diaspora Populations Comprise Distinct Genetic Clusters with Shared Middle Eastern Ancestry. The American Journal of Human Genetics, 03 June 2010. http://www.cell.com/AJHG/abstract/S0002-9297%2810%2900246-6 Cell http://www.cell.com/
- Behar D.M., Thomas M.G., Skorecki K., Hammer M.F., Bulygina E., Rosengarten D., Jones A.L., Held K., Moses V., Goldstein D., Bradman N., Weale M.E. Multiple origins of Ashkenazi Levites: Y chromosome evidence for both Near Eastern and European ancestries. Am J Hum Genet. 2003 Oct;73(4):768-79. Epub 2003 Sep 17. http://www.familytreedna.com/pdf/400971.pdf Family Tree DNA. http://www.familytreedna.com/
Behar D.M., Yunusbayev B., Metspalu M., Metspalu E., Rosset S., Parik J., Rootsi S., Chaubey G., Kutuev I., Yudkovsky G., Khusnutdinova E.K., Balanovsky O., Semino O., Pereira L., Comas D., Gurwitz D., Bonne-Tamir B, Parfitt T., Hammer M.F., Skorecki K., Villems R. The genome-wide structure of the Jewish people. Nature. 2010. http://www.nature.com/nature/journal/vaop/ncurrent/full/nature09103.html nature.com http://www.nature.com/ - Hammer M.F., Behar D.M., Karafet T.M., Mendez F.L.,
Hallmark B., Erez T., Zhivotovsky L.A., Rosset S., Skorecki K. Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood. Hum Genet. Received: 8 April 2009 / Accepted: 25 July 2009. http://www.springerlink.com/content/357176p177623m41/fulltext.pdf This article is published with open access at Springerlink.com - Zoossmann-Diskin A. The origin of Eastern European Jews revealed by autosomal, sex chromosomal and mtDNA polymorphisms. http://www.ncbi.nlm.nih.gov/pubmed/20925954?dopt=Abstract Biol Direct. 2010 Oct 6;5:57.
