Генохронологическое датирование евреев гаплогруппы E1b1b1
Тюрин Анатолий Матвеевич, к.г.-м.н.
Ключевые слова: евреи, популяционная генетика, генохронология, гаплогруппа E1b1b1(M35).
Оглавление
1. Введение
2. Особенности генетического портрета евреев
3. Результаты генохронологического датирования
4. География линий E1b1b1*, E1b1b1a3*, E1b1b1c1* и E1b1b1c1a
5. Интерпретация результатов датирования
6. Проблемы генохронологического датирования
7. Вместо заключения
Источники информации
Результаты генохронологического датирования кластеров гаплогруппы E1b1b1 в табличном виде
1. Введение
Автор публикации [Алиев, 2010] выполнил по базе данных E3b Cluster генохронологическое датирование общих предков (TMRCA) кластеров гаплотипов линий гаплогруппы E1b1b1(M35) Y-хромосомы. Результаты характеризуют евреев ашкенази по мужской линии. Датирование выполнено по 37 и 67 маркерным гаплотипам. Для четырех кластеров гаплотипов получены даты жизни их общих предков – 9-11 века, для других двух кластеров – 2-3 века н.э. Эти даты увязаны с историей евреев. Однако сегодня пока не решены некоторые важные методические вопросы генохронологического датирования. Главный из них – достоверное выделение кластеров гаплотипов, восходящих к общим предкам. В публикации [Алиев, 2010] принципы выделения датируемых кластеров гаплотипов и их структура не рассмотрены. Очевидно, ее автор считает, что в этом вопросе никаких проблем нет и, следовательно, техническую часть генохронологического датирования можно в статье не приводить. Достаточно дать ссылку на прекрасно организованную базу данных E3b Cluster и любой, владеющий основами генохронологического датирования может выполнить его сам. Нам представлялось интересным это сделать.
2. Особенности генетического портрета евреев
Массив ДНК-данных по евреям приведен в публикации [Behar, 2003]. Гаплогруппы имеют невысокое разрешение. Выборка включает коэнов ашкенази (76 человек), левитов ашкенази (60 человек), израэлитов ашкенази (100 человек), коэнов сефардов (69 человек), левитов сефардов (31 человек) и израэлитов сефардов (63 человека). У них доминируют носители гаплогруппы J: 86,8%, 10,0%, 37,0%, 75,4%, 32,3%, 36,5%. Носители гаплогруппы E*(xE3a) среди евреев составляют 4,0%, 20,0%, 22,0%, 4,4%, 9,7% и 19,1%. Кроме того для сопоставления приведены результаты ДНК-тестирования немцев (выборка 88 человек), норвежцев (83 человека), лужицких сербов (112 человек) и белорусов (360 человек). У них носители гаплогруппы E*(xE3a) составляют 3,4%, 1,2%, 6,3% и 4,6%. Эти данные дают только самые общие представления о географии гаплогруппы Е.
Высоко разрешенные ДНК-данные по евреям приведены в публикации [Hammer, 2009]. Они включают гаплогруппы и гаплотипы 1575 евреев, в том числе 215 коэнов, 154 левитов, 738 израэлитов и 468, не знающих к какой из трех каст они относятся. Данные по частотам гаплогрупп в табличной форме приведены только для коэнов и израэлитов без разделения их на ашкенази и не ашкенази. Носителей гаплогруппы E1b1b1(M35) среди коэнов не выявлено. Среди израэлитов их 20 (2,7%). Всего носителей гаплогруппы Е среди коэнов – 15 (7,0%), среди израэлитов – 138 (18,7%). Носителей гаплогруппы J среди коэнов – 162 (75,3%), израэлитов – 265 (35,9%).
Авторы публикации [Adams, 2008] выполнили сопоставление генетического портретов сефардов (выборка 174 человека), жителей Пиренейского полуострова (1140 человек) и жителей стран Магриба (361 человек). Среди сефардов 46,6% носителей гаплогруппы J и 9,2% носителей гапогруппы E. Но носителей линии E1 среди них не выявлено.
Таким образом, результаты датирования евреев гаплогруппы E1b1b1 никак не характеризуют этапы формирования жреческой касты коэнов и сефардов Испании. Среди них нет носителей этой гаплогруппы. Относительно другой жреческой касты – левитов, данных у нас нет. Зато можно однозначно утверждать, что эти результаты характеризуют этапы формирования израэлитов, самой многочисленной касты евреев.
3. Результаты генохронологического датирования
Генохронологическое датирование евреев гаплогруппы E1b1b1 выполнено исходя из нашего понимания принципов выделения кластеров гаплотипов, восходящих к общему предку. При этом во внимание приняты 12 маркерные гаплотипы (DYS393, DYS390, DYS19, DYS391, DYS385a, DYS385b, DYS426, DYS388, DYS439, DYS389-1, DYS392, DYS389-2). Это существенно расширяет статистическую базу датирования, по сравнению с датированием по 37 и 67 маркерным гаплотипам, что позволяет лучше видеть структуру выборок. Скорость мутаций принята равной 0,002 на маркер на поколение [Тюрин, 2009, Скорости мутаций]. «Поколение» равно 25 годам. За расчетный принят 1980 год (при допущении, что средний возраст тестированных примерно равен 30 годам).
Выделенные в группах гаплотипов кластеры линий E1b1b1*, E1b1b1a3*, E1b1b1c1* и E1b1b1c1a приведены в таблицах 1-6. Сводные результаты датирования – в таблице 7. Выделено 14 кластеров гаплотипов, 13 из них датированы. Всего при датировании евреев принято во внимание 240 гаплотипов. Автор публикации [Алиев, 2010] принял во внимание 101 гаплотип. Результаты датирования пересчитаны в параметр, характеризующий количество евреев, датированные предки которых жили в конкретном веке (рисунок 1). При его расчете количество гаплотипов кластеров, дата общего предка которых попала на рубеж веков (даты «1500 год» и «1600 год»), делилось в равных пропорциях между сопряженными веками.
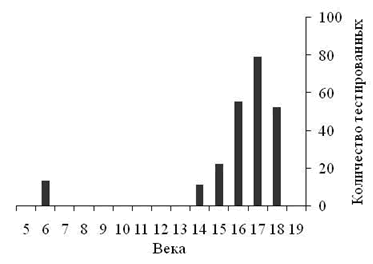
Рисунок 1. Сводные результаты генохронологического датирования евреев гаплогруппы E1b1b1.
Ось ординат характеризует количество евреев, датированные предки которых жили в конкретном веке.
4. География линий E1b1b1*, E1b1b1a3*, E1b1b1c1* и E1b1b1c1a
В базе данных E3b Cluster для всех тестированных евреев указаны фамилии и территория проживания их ближайших предков (origin). Предки евреев компактно сгруппировались на территории Украины – 39, Польши – 30, России – 23, Белоруссии – 21, Литвы – 20, и Германии – 18. Всего 154 (61,2%). Другие европейские страны указали 38 (15,8%) тестированных. Три из них указали Латинскую Америку, два – Израиль (?). 43 (17,9%) евреев не указали место жительства своих предков. По этим данным можно сделать следующие выводы. Предки наибольшего числа тестированных евреев проживали в регионе расселения на рубеже 19-20 веков ашкенази. Последних и характеризует рассматриваемая выборка гаплотипов. Такой же вывод, но по фамилиям тестированных, сделал так же автор публикации [Алиев, 2010]. Кроме того, напомним, что среди сефардов Испании не выявлены носители линии E1.
В гаплотипах базы данных E3b Cluster в линиях E1b1b1*, E1b1b1a3*, E1b1b1c1* и E1b1b1c1a выделены и «не еврейские» группы. Их география принципиально отличается от географии еврейских групп. Исключение составляет группа E1b1b1c1a-B. Приведенные ниже статистические выкладки ее не учитывают. Всего рассматриваемые группы включают 223 гаплотипа. Территорию расселения ашкенази, как место жительства своих предков, указали 24 (10,8%) тестированных не евреев, в том числе, 20 – Германию и по 1 – Польшу, Россию, Белоруссию и Украину. Другие европейские страны указали 110 (49,3%) тестированных. Ближний Восток – 15, другие страны – Азии и Африки – 12, страны Американского континента – 9. Не указали родину своих предков – 77 (34,5%) тестированных. Зафиксируем еще одну цифру. Территорию Греции, Турции и Кипра как место проживания своих предков, указали 7 тестированных не евреев. То есть Восточное Средиземноморье и Ближний Восток указало 22 (9,9%) не евреев. Эта цифра плохо согласуются с представлениями о том, что эти регионы являлись местом первоначального расселения носителей гаплогруппы E1b1b1 [Алиев, 2010].
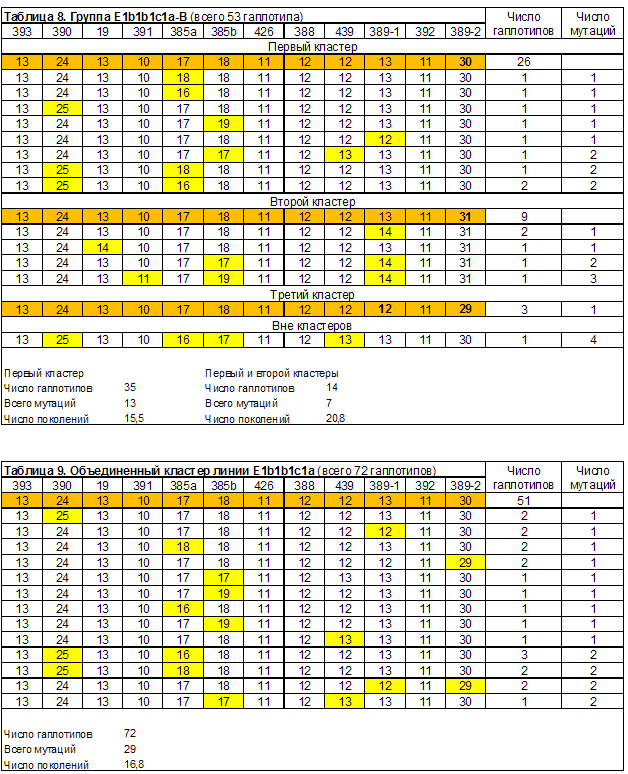
Предки тестированных не евреев, попавших в группу E1b1b1c1a-B (всего 53 гаплотипа), компактно сгруппировались на той же территории, что и предки евреев: Украина – 12, Польша – 5, Россия – 2, Белоруссия – 6, Литва – 4, и Германия – 6. Всего 33 (66,0%). По 3 тестированных указали, как место жительства их предков, Словакию и Израиль (?), по 1 – Голландию и Англию. 9 тестированных не указали место жительства предков. Если мы исключим из групп евреев тех, кто не указал место жительства своих предков, то получим, что 78,2% из них родом с территории проживания ашкенази. Для нееврейской группы E1b1b1c1a-B эта цифра составляет 75,0%.
Датирование общих предков двух кластеров группы E1b1b1c1a-B (таблица 8) дало те же цифры, что и для большинства еврейках кластеров. У этой группы имеется яркая особенность. Ее первый кластер совпадает с первым кластером группы E1b1b1c1a-A. Совпадают модальные и 5 связанный с ними гаплотипов. Это дает возможность объединить два указанных кластера (таблица 9). Датирование объединенного кластера линии E1b1b1c1a (всего 72 гаплотипа) дало время жизни общего предка 16,8 поколений или 420 лет назад. Это 1560 год. Отметим высокую сходимость результатов оценки жизни общего предка, двух кластеров, сформированных по национальному признаку на основе территориально обособившегося объединенного кластера. Для евреев это 18,0, для не евреев – 15,5 поколений назад. Отметим, что совпадение гаплотипа, который мы не включили в кластеры группы E1b1b1c1a-B (таблица 8), с одним из аналогичных гаплотипов группы E1b1b1c1a-A (таблица 5), связано с погрешностью формирования выборок. Это результаты тестирования одного и того же человека (фамилия – Caligur, номер теста – 71213, место жительства предков не указано).
5. Интерпретация результатов датирования
Только по информации, опубликованной в базе данных E3b Cluster без привлечения каких либо априорных сведений, получены вполне достоверные результаты.
1. Датированы общие предки евреев ашкенази, носители линий E1b1b1*, E1b1b1a3*, E1b1b1c1* и E1b1b1c1a. Общие предки большинства тестированных жили в 14-18 веках.
2. Предки не евреев этих линий жили, в основном, за пределами исторического региона расселения ашкенази.
3. Один из кластеров не евреев группы E1b1b1c1a-B идентичен кластеру евреев группы E1b1b1c1a-A.
Вместе с тем, по результатам анализа базы данных не удалось локализовать географический регион первоначального проживания носителей гаплогруппы E1b1b1. Автор публикации [Алиев, 2010] на основе современной геногеографии локализует его в Восточном Средиземноморье и на Ближнем Востоке. При учете результатов нашего датирования мы конкретизируем эту локализацию. Регионом первоначального проживания носителей рассматриваемой гаплогруппы была Византийская, а затем Османская империи. Теперь у нас есть все, для выполнения популяционной реконструкции. Результат датирования общего предка второго кластера группы E1b1b1*-D (таблица 2) – 57,7 поколений или 1445 лет назад, мы пока рассматривать не будем, поскольку не выполнен углубленный анализ (на основе формальных процедур обработки ДНК-данных) его достоверности. Такой анализ необходим именно потому, что дата общего предка кластера кардинально не соответствует другим 11 датам.
Начиная с 14 века с территории Византии, а затем Османской Турции существовало перманентное переселение некоторых социальных групп на территорию Европы за пределы Балкан. Среди них были и носители линий E1b1b1*, E1b1b1a3*, E1b1b1c1* и E1b1b1c1a. Они явились предками людей, гаплотипы которых сформировали датированные кластеры. Но здесь имеется один принципиальный момент. География линий гаплогруппы E1b1b1 такова, что вариант формирования кластеров от предков на территории Европы представляется маловероятным. Первичное формирование кластеров произошло на «родине» гаплогруппы E1b1b1. На территорию Европы переселялись представители социальных групп, среди которых уже существовали носители обособившихся кластеров гаплотипов. Те из них, которые попадали в регион формирования религиозно-этнической общности ашкенази, пополняли их общины. Это привело к тому, что потомки мигрантов, в своем большинстве, являются сегодня евреями, а среди коренного населения региона, их потомков относительно немного. Те мигранты, которые попадали за пределы региона формирования ашкенази, ассимилировались среди местного населения. Поэтому среди коренных жителей Европы за пределами обозначенного региона носителей рассматриваемых линий заметно больше, чем среди проживающих в нем. У носителей гаплотипов объединенного кластера линии E1b1b1c1a особая история. Либо их предки примерно поровну вливались в еврейские и не еврейские социальные сообщества, либо не евреи носители линии E1b1b1c1a являются потомками евреев. Против второй версии свидетельствует относительно небольшое число носителей других рассматриваемых линий среди не евреев на территории проживания ашкенази.
Выскажем и вторую версию интерпретации полученных результатов. Большинство не евреев гаплогруппы E1b1b1 являются потомками евреев. В регионе проживания ашкенази евреи были жестко обособлены от его других социальных сообществ территориально, культурно и по исповедуемой ими религии. Поэтому здесь процессы их ассимиляции не имели значительного размаха. При иммиграции евреев за пределы региона проживания ашкенази их потомки в течение первых поколений ассимилировались среди коренного населения. Это и отражают особенности географии гаплогруппы E1b1b1
6. Проблемы генохронологического датирования
Ненормальное положение дел в области оценок скоростей мутаций маркеров гаплотипов Y-хромосомы и их применения в датировании, является, по нашему мнению, следствием сознательного запутывания этого вопроса [Тюрин, 2009, Скорости мутаций]. Но другой вопрос – выделение кластеров гаплотипов, восходящих к одному предку, не просто запутан, но находится в зоне «молчания». Его цена показана на примере датирования якутов [Тюрин, 2010, Генохронология якутов]. Попросту говоря, мы утверждаем, что авторы публикаций [Адамов, 2008; Харьков, 2008; Pakendorf, 2006] выполнили некорректное генохронологическое датирование общего предка якутов гаплогруппы N1c1. У якутов, носителей этой линии, не один модальный гаплотип, как считают авторы публикаций, а три. То есть, якуты, носители гаплогруппы N1c1 восходят к трем общим предкам. На этом и должно базироваться их генохронологическое датирование.
Проблема оценки корректности выделения кластеров «коротких» (не более 17-25-ти маркеров) гаплотипов решается предельно просто. Необходимо создать имитационную модель, на которой можно будет просчитывать прямые задачи генохронологического датирования. На вход модели должно подаваться всего три параметра, характеризующих датированный кластер: количество маркеров в гаплотипах, принятая скорость мутаций и собственно результат датирования. Результатами моделирования должно быть относительное распределение количества разных гаплотипов (модального и отличных от него на 1 и более шагов), а также вероятные величины одинаковых гаплотипов. В создании такой модели имеется только одна проблема. Выполненные на ней расчеты «опрокинут» результаты генохронологического датирования очень многих авторов. Отметим, что автор публикации [Адамов, 2008] при датировании якутов выполнил теоретический расчет распределения количества гаплотипов. Но расчет вероятных величин одинаковых гаплотипов он не произвел.
В выделении кластеров гаплотипов имеется еще одна проблема. В гаплотипах группы E1b1b1c1*-D1 мы выделили четыре кластера, каждый из которых связан с одним из модальных гаплотипов (таблица 4). Структура выборки проста и понятна. На ее основе выдвигается конкретная гипотеза: каждый кластер связан с одним предком, который имел конкретный 12 маркерный гаплотип. Это и является основанием генохронологического датирования. То есть, само предположение о наличии общего предка (мы говорим о ближайшем общем предке, который и датируется генохронологическим методом) у людей, гаплотипы которых включены в кластер и имеют определенную длину, является фикцией. На самом деле количество маркеров в Y-хромосомы, отражающее родство людей одной линии гаплогруппы, на порядки превышает, например, те 12 маркеров, которыми мы ограничились. Это легко понять при рассмотрении 67 маркерных гаплотипов группы E1b1b1c1*-D1. Среди них нет ни одного гаплотипа, который мог бы быть принят за модальный. По крайней мере, мы такого не нашли. Автор публикации [Алиев, 2010], как мы понимаем, смог датировать эту группу только на основе суррогатного модального гаплотипа. Это и есть гаплотип фиктивного предка. Нам представляется, что применение при генохронологическом датировании «фиктивного предка» является вполне корректной операцией. Только нужно четко понимать, что общий предок кластера гаплотипов фиктивен вне зависимости от числа маркеров в гаплотипах кластера. Можно датировать кластер и по одному маркеру. Можно датировать по 6, 12, …. 67, ….. маркерным гаплотипам. Это не принципиально. Принципиально то, что общий предок всегда фиктивен, поскольку датируется не он, но время возникновения участка Y-хромосомы определенной длины. Случаи с реальными предками групп людей рассматриваются в рамках другой дисциплины популяционной генетики – геногенеалогии.
Теперь можно сравнить результаты датирования группы E1b1b1c1*-D1. Автор публикации [Алиев, 2010] принял ее за единый кластер. Получена дата общего предка – 40,0 поколений или 1000 лет назад. Мы выделили в этой группе 4 кластера (таблица 4). Даты их предков – 9,6, 11,3, 26,5, 13,9 поколений назад. Если объединить второй кластер с третьим, а первый с четвертым, то даты предков будет 25,2 и 15,7 поколений назад. Результаты двух датирований различаются в разы. Мы выполнили датирование по 12 маркерным гаплотипам, автор публикации [Алиев, 2010] – по 67. Но нашему датированию предшествовал детальный анализ структуры выборки с выделением в ней кластеров. А какова структура 67 маркерных гаплотипов группы E1b1b1c1*-D1?
При анализе структура 67 маркерных гаплотипов (всего 34) группы E1b1b1c1*-D1 мы применили два способа. Первый из них – выделение мод в числе аллель каждого маркера гаплотипа, второй – трассирование модального гаплотипа. Маркеры гаплотипов, не имеющие мутаций, из анализа исключены. Всего осталось 27 маркеров из 67 (рисунок 2). В 6 маркерах четко обозначилось две моды, в остальных – одна мода. Суммарное количество мутаций в маркерах с одной модой – 41 в сторону уменьшения числа аллель и 38 в сторону его увеличения. Хорошее соответствие этих цифр говорит о том, что мутации в маркерах с одной модой являются случайным процессом с нормальным распределением его результатов. Две моды в маркере свидетельствует о том, что в выборку включены гаплотипы, восходящие к разным общим предкам. В числе аллель маркера DYS389-2 четко обозначилось две моды – 30 и 31. Можно скомпоновать соответствующие им гаплотипы в две выборки и снова выделить в них моды (рисунки 3 и 4). В выборке DYS389-2 = 31 5 маркеров имеют две моды в числе аллель, в выборке DYS389-2 = 31 – 1 маркер. В выборке 9 маркерных гапотипов якутов (172 гаплотипа) [Pakendorf, 2006] маркеры DYS392 и DYS389-2 имеют две моды числа аллель (рисунок 5). То есть эта выборке не может быть отнесена к одному общему предку, что мы и обосновали в публикации [Тюрин, 2010, Генохронология якутов].
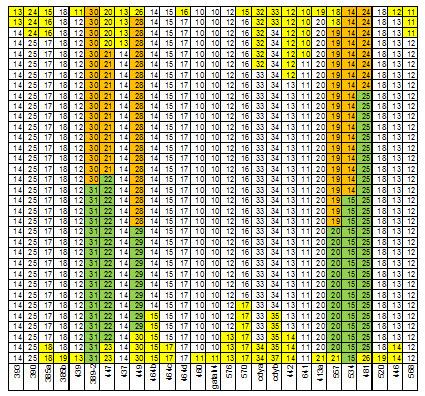
Рисунок 2. Структура 67 маркерных гаплотипов группы E1b1b1c1*-D1.
Маркеры, имеющие только одно значение количества аллель, не показаны.
Желтым цветом отмечены маркеры, число аллель которых отличается от его модального значения.
Зеленым и коричневым цветами показаны две разные моды в числе аллель маркера.
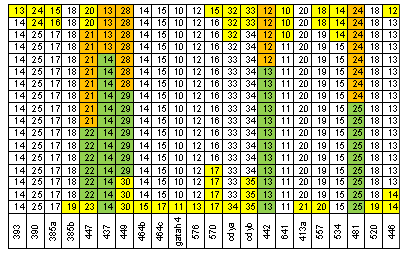
Рисунок 3. Структура 67 маркерных гаплотипов группы E1b1b1c1*-D1 для моды DYS389-2 = 31.
Пояснения под рисунком 2.
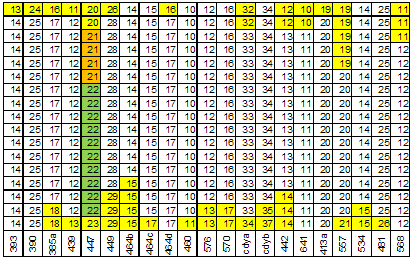
Рисунок 4. Структура 67 маркерных гаплотипов группы E1b1b1c1*-D1 для моды DYS389-2 = 30.
Пояснения под рисунком 2.
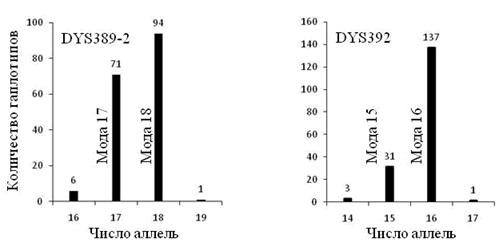
Рисунок 5. Структура 9 маркерных гаплотипов якутов [Pakendorf, 2006].
Распределение числа аллель в маркерах DYS392 и DYS389-2.
Идея трассирования модального гаплотипа проста. В группе E1b1b1c1*-D1 67 маркерных гаплотипов можно обособить, например, 12 первых из них и для них выделить модальный гаплотип. Это сделал автор публикации [Алиев, 2010] – 14, 25, 13, 9, 17, 18, 11, 12, 12, 13, 11, 30. Таких 17 гаплотипов. Что с ними происходит при увеличении количества маркеров в гаплотипе? Ответ приведен на рисунке 6. Если мы примем во внимание 32 маркера, то в выборке останется 3 модальных гаплотипа. Если 33 – то модального гаплотипа в ней не будет. Один из экземпляров модального гаплотипа «пронизывает» модальные значения числа аллель всех 67 маркеров. Если во внимание примем 12 последних маркеров, то среди них тоже выделяется модальный гаплотип, представленный 8 экземплярами. На этом примере хорошо видно, то, что общий предок кластера гаплотипов – фикция. Похоже, это не понимают те, кто пытается увязать 6 и 12 маркерные модальные гаплотипы коэнов с библейским Авраамом. Но вернемся к рисунку 6. На нем хорошо видно, что на маркере DYS447 «прерываются» 7 из 14 модальных гапотипов. Здесь имеется две возможности. Первая – маркер DYS447 имеет высокую скорость мутаций, вторая – число аллель в нем имеет более чем 1 моду. Но возможно справедливы оба предположения. Для нас важно то, что этот маркер действительно имеет две моды – 21 и 22 (рисунок 2-4). Здесь имеется еще один интересный момент. Три модальных гаплотипа «утыкаются» в значения 20 и 23 числа аллель маркера DYS447, то есть в маркеры, число аллель которых отличается от модальных значений. Это означает, что три гаплотипа попали в кластер модальных 12 маркерных гаплотипов случайно. В общем-то, все 17 модальных 12 маркерных гаплотипов сгруппировались в выборке группе E1b1b1c1*-D1 случайно. Именно случайный характер этого явления и позволяет считать вполне корректным введение гипотезы об их отнесении к единому фиктивному общему предку.
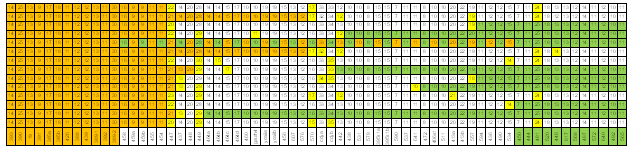
Рисунок 6. Структура 67 маркерных гаплотипов группы E1b1b1c1*-D1. Трассирование 12 маркерных модальных гаплотипов. Желтым цветом отмечены маркеры, число аллель которых отличается от его модального значения. Зеленым и коричневым цветами показаны модальные гаплотипы для первых и последних 12 маркеров.
Наш общий вывод однозначен. Группа гаплотипов E1b1b1c1*-D1 не может быть отнесена к одному общему предку и, следовательно, последний не может быть датирован формальным генохронологическим методом. Отметим, что формальное датирование ее предка даст системно завышенный результат, который и получил автор публикации [Алиев, 2010].
7. Вместо заключения
В публикации [Алиев, 2010] техническая часть расчетов ограничена следующей фразой: «По алгоритму А.А. Клёсова [13] произведен расчет возрастов ближайших общих предков …». Смотрим «Руководство к расчету времен до общего предка гаплотипов Y-хромосомы …» [Клёсов, 2008]. В нем четко обозначен первый шаг генохронологического датирования. «Первое – убедиться, что для рассматриваемой выборки гаплотипов есть всего один предок, или соответствующий ему эквивалент. Эквивалентом могут быть братья или другие близкие родственники (что уже находится вне предела разрешения методами ДНК-генеалогии). … Для того, чтобы убедиться, что выборка «однородная», то есть происходит от одного общего предка, есть несколько приемов.». Мы дадим другую редакцию первого шага генохронологического датирования. Необходимо выполнить анализ структуры датируемой выборки гаплотипов с выделением в ней их кластеров, восходящих к одному предку. По результатам анализа принимается решение о возможности корректного датирования. В случае публикации результатов датирования, в статье должны быть приведены все обоснования правомочности его применения.
Алиев А. О происхождении «еврейских» кластеров гаплогруппы E1b1b1 (M35). The Russian Journal of Genetic Genealogy (Русская версия), Vol 2, No 1 (2010). http://rjgg.molgen.org/index.php/RJGGRE Popular Science Journals. http://rjgg.molgen.org/
Клёсов А. Руководство к расчету времен до общего предка гаплотипов Y-хромосомы и таблица возвратных мутаций. Вестник Российской Академии
ДНК-генеалогии. Том 1, № 5. 2008. Michael Temosh's Storefront. http://stores.lulu.com/temosh
[Тюрин, 2010, Скорости мутаций] Тюрин А.М. «Трансмиссионные», «эволюционные» и «сконструированные» скорости мутаций локусов гаплотипов Y-хромосомы. /volume10/turin_mutation.php. Электронный сборник статей «Новая Хронология». Выпуск 10. 2010. /volume10/ Сайт: Новая Хронология. http://www.chronologia.org/
[Тюрин, 2010, Генохронология якутов] Тюрин А.М. К вопросу о генохронологическом датировании предков якутов. /volume10/turin_jakuty.php. Электронный сборник статей «Новая Хронология». Выпуск 10. 2010. /volume10/ Сайт: Новая Хронология. http://www.chronologia.org/
Харьков В.Н. Степанов В.А., Медведева О.Ф., Спиридонова М.Г., Максимова Н.Р., Ноговицына А.Н., Пузырев В.П. Происхождение якутов: анализ гаплотипов Y-хромосомы. Молекулярная биология, 2008, том 42, № 2, с.226-237. http://www.maikonline.com/maik/showArticle.do?auid=VAFDZK2NOW&lang=ru Электронные версии научных журналов. http://www.maikonline.com/
Adams S.M., Bosch E., Balaresque P.L., Ballereau S.J., Lee A.C., Arroyo E., Lopez-Parra A.M., Aler M., Grifo M.S.G., Brion M., Carracedo A., Lavinha J., Martinez-Jarreta B., Quintana-Murci L., Picornell A., Ramon M., Skorecki K., Behar D.M., Calafell F. and Jobling M.A. The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula. The American Journal of Human Genetics, Volume 83, Issue 6, 725-736, 04 December 2008.
https://lra.le.ac.uk/bitstream/2381/4116/1/Adams08.AJHG_LRA.pdf University of Leicester. https://lra.le.ac.uk/
Behar D.M., Thomas M.G., Skorecki K., Hammer M.F., Bulygina E., Rosengarten D., Jones A.L., Held K., Moses V., Goldstein D., Bradman N., Weale M.E. Multiple origins of Ashkenazi Levites: Y chromosome evidence for both Near Eastern and European ancestries. Am J Hum Genet. 2003 Oct;73(4):768-79. Epub 2003 Sep 17. http://www.familytreedna.com/pdf/400971.pdf Family Tree DNA. http://www.familytreedna.com/
Hammer M.F., Behar D.M., Karafet T.M., Mendez F.L.,
Hallmark B., Erez T., Zhivotovsky L.A., Rosset S., Skorecki K. Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood. Hum Genet. Received: 8 April 2009 / Accepted: 25 July 2009. http://www.springerlink.com/content/357176p177623m41/fulltext.pdf This article is published with open access at Springerlink.com
Pakendorf B., Novgorodov I.N., Osakovskij V.L., Danilova A.P., Protod'jakonov A.P., Stoneking M. Investigating the effects of prehistoric migrations in Siberia: genetic variation and the origins of Yakuts. Hum.Genet., 2006, 120: 334-353. http://www.eva.mpg.de/genetics/pdf/Yakut_article_2006.pdf
The Max Planck Institute for Evolutionary Anthropology. http://www.eva.mpg.de/
Результаты генохронологического датирования
кластеров гаплогруппы E1b1b1 в табличном виде
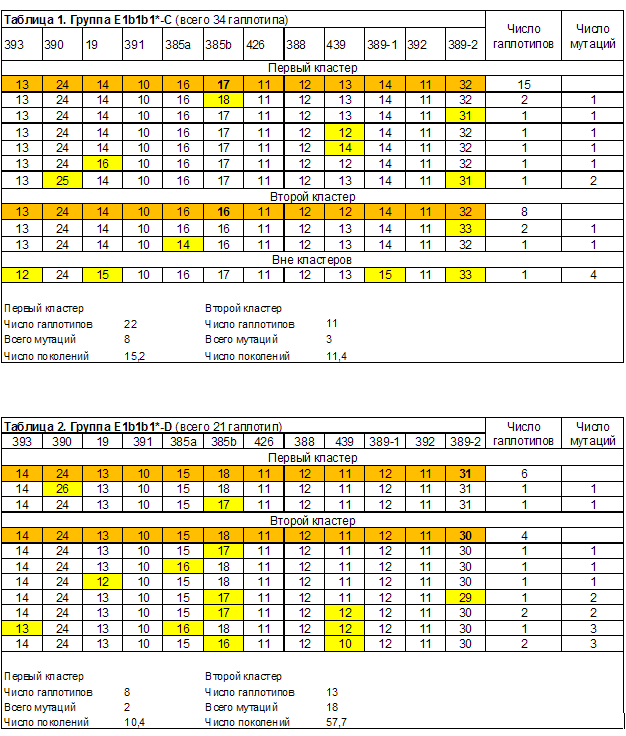
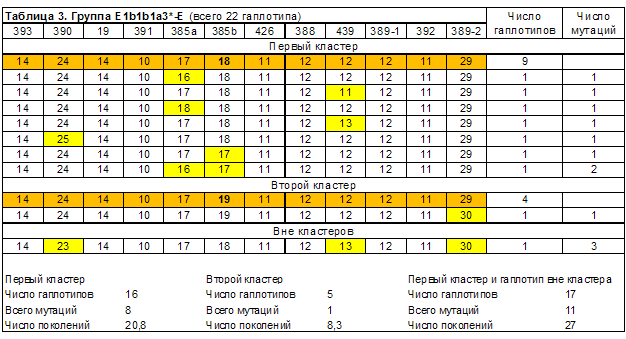
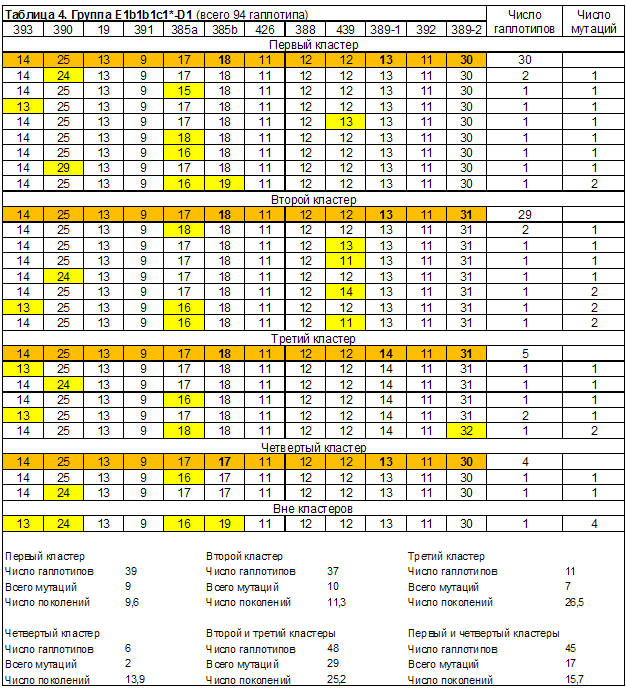
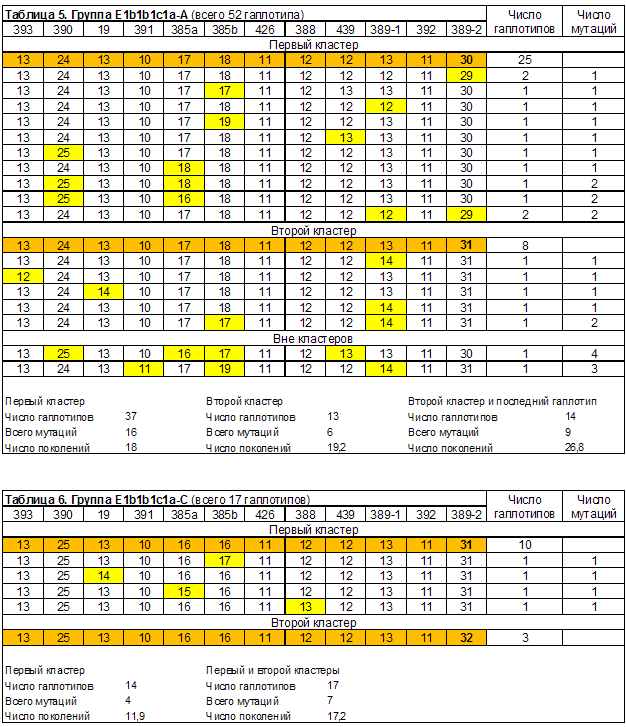
Таблица 7.
Сводные результаты генохронологического датирования кластеров гаплогруппы E1b1b1.
| Кластеры | Число гаплотипов | Время жизни общего предка | ||
| Поколений назад | Лет назад | Календарный год | ||
| Группа E1b1b1*-C. Всего 34 гаплотипа. | ||||
| Первый кластер | 22 | 15,2 | 380 | 1600 |
| Второй кластер | 11 | 11,4 | 285 | 1695 |
| Группа E1b1b1*-D. Всего 21 гаплотип. | ||||
| Первый кластер | 8 | 10,4 | 260 | 1720 |
| Второй кластер | 13 | 57,7 | 1445 | 535 |
| Группа E1b1b1a3*-E. Всего 22 гаплотипа. | ||||
| Первый кластер | 16 | 20,8 | 520 | 1460 |
| Второй кластер | 5 | 8,3 | 210 | 1770 |
| Группа E1b1b1c1*-D1. Всего 94 гаплотипа. | ||||
| Первый кластер | 39 | 9,6 | 240 | 1740 |
| Второй кластер | 37 | 11,3 | 285 | 1695 |
| Третий кластер | 11 | 26,5 | 665 | 1315 |
| Четвертый кластер | 6 | 13,9 | 350 | 1630 |
| Группа E1b1b1c1a-A. Всего 52 гаплотипа. | ||||
| Первый кластер | 37 | 18 | 450 | 1530 |
| Второй кластер | 13 | 19,2 | 480 | 1500 |
| Группа E1b1b1c1a-C. Всего 17 гаплотипов. | ||||
| Первый кластер | 14 | 11,9 | 300 | 1680 |
| Группа E1b1b1c1a-B. Всего 53 гаплотипа. | ||||
| Первый кластер | 35 | 15,5 | 390 | 1590 |
| Второй кластер | 14 | 20,8 | 520 | 1460 |
| Линия E1b1b1c1a. | ||||
| Объединенный кластер | 72 | 16,8 | 420 | 1560 |